


成行成田:通过调控作物结构的冬小麦田展现出生态集约化潜力——基于更强的天然害虫与杂草种子控制能力
发布时间:2025-08-29 13:36 | 点击次数:184
成行成田:通过调控作物结构的冬小麦田展现出生态集约化潜力——基于更强的天然害虫与杂草种子控制能力
原文题目:Rows make the field: Winter wheat fields with manipulated crop architecture show potential for ecological intensification based on higher natural pest and weed seed control.

亮点
• 调控作物结构可作为田间生态集约化的创新措施。
• 通过简单关闭部分播种开沟器即可实现,无需减少播种量。
• 不同作物密度下的结构调控田块具有更高的生境异质性。
• 步甲群落的变化与害虫控制服务呈正相关。
• 具备在不降低小麦产量的前提下减少农药使用的潜力。
摘要
生态集约化旨在通过增加生物多样性及利用相关生态系统服务,来替代或结合外部投入,从而降低农业生产的环境成本,同时维持或提高作物产量。然而,目前能在田间实际验证该效益的措施仍十分有限。
本研究通过调控冬小麦田的播种模式,在试验田构建由"未播种行"和"密播行"组成的多样化作物结构,与对照田的"常规播种行"进行比较,以评估:(i) 调控作物结构的田块是否具有更异质的田间结构和小气候条件;(ii) 作物结构改变是否会提高步甲群落物种丰富度、活动密度并改变体型分布;(iii) 这是否会增强天然害虫与杂草种子控制能力;(iv) 且不会导致产量损失。
结果表明:试验田的田间结构和小气候条件比对照田更具异质性,这与捕食性步甲体型显著增大及物种丰富度增加趋势(出现于试验田"未播种行")相关。这些变化使试验田"密播行"的害虫抑制能力显著优于对照田"常规播种行"——实验害虫捕食率提高10%,谷物叶甲造成的作物损害减少56%。此外,"未播种行"中已知取食杂草种子的步甲活动密度显著高于"常规播种行",这与"密播行"实验杂草种子捕食率显著升高相关。调控作物结构与否的田块小麦产量无显著差异。
研究表明:通过简单调整冬小麦播种模式创建多样化作物结构,可促进步甲多样性,改变其群落组成和活动密度,从而在不造成减产的前提下增强关键生态系统服务。因此,该措施除保护生物多样性外,虽未直接验证,但具备通过增强生态系统服务实现农药生态替代的潜力,且不会产生土地机会成本或增加工作量等农艺与经济劣势。
关键词
生态替代 | 生境管理 | 谷物叶甲属 | 扩散后种子捕食 | 作物密度 | 天敌
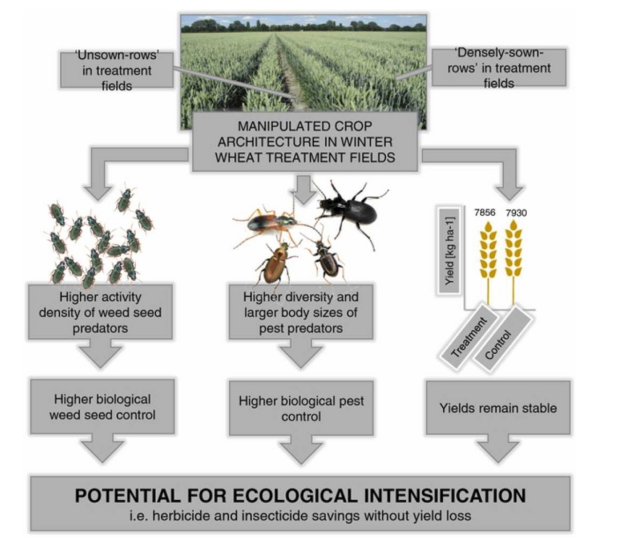
生态集约化潜力—节省除草剂和杀虫剂而不损失产量
引言
集约化农业正全球性威胁农田生物多样性及相关生态系统服务(Foley等,2005)。主要驱动因素包括化肥、农药等外部投入,这些虽能增产或避免减产,但对生物多样性有害(Tilman等,2001;Kleijn等,2019)。例如,农药已导致非靶标甲虫(Lee等,2001)、蜜蜂(Alston等,2007)和鸟类(Hart等,2006)数量下降。
作物主导区尤其具有高农药投入特征(Hossard等,2014),其中冬小麦田在欧洲占比超25%(基于2021年FAO统计数据)。因此,在此类区域开发农药的可持续替代方案是当前重大挑战,欧盟绿色协议也提出农药减量目标(EU Commission,2019)。特别困难的是如何在保证作物总产不降的前提下实现该目标,因为农产品需求持续增长(Glamann等,2017)。有机农业虽能减轻环境负面影响,但通常伴随减产,需更多土地生产等量作物,这可能对生物多样性产生负面效应,因而不足以完全解决这些问题(Tscharntke等,2021)。
相比之下,生态集约化力求最小化环境负面影响,同时满足农田产品增长需求(Bommarco等,2013)。该理念旨在用生物多样性提供的服务替代或补充外部投入,如通过天敌天然控害替代农药。基于"可通过管理特定生物多样性组分进一步提升产量(生态增效)或替代外部投入而不负面影响生产(生态替代)"的假设,生态集约化致力于降低农业生产的环境成本(如对生物多样性的负面影响)(Kleijn等,2019)。尽管相关研究日益增多,但目前仍缺乏有效且易实施的生态集约化措施,其生物多样性及关联生态系统服务的效益尚未经田间验证(Bommarco等,2013;Lyu等,2021)。包含产量评估并考量措施对农民需求的研究更少,导致大规模推广困难。尤其在小麦生产体系中,能促进生物多样性及天然控害服务、且不产生显著农艺或经济成本的成功案例仍稀缺(但见Tschumi等,2016;Pywell等,2015)。
作物结构调控(即通过田间不同作物密度设计特定播种模式)可能是此类管理选项之一,可增加作物结构多样性。根据结构异质性假说(Tews等,2004),作物结构多样性增加可能对节肢动物等高营养级多样性产生上行效应,从而提升害虫控制或杂草种子捕食等生态系统服务(Kromp,1999;Menalled等,2006)。具有更高结构异质性和多样小气候条件的田块,可能更好满足有益节肢动物的物种特异性生境偏好,进而改变功能性状(如体型)组成,并通过更大体型天敌的单位个体更高害虫消耗量等途径,对控害服务产生积极影响(Rouabah等,2014)。然而,关于作物结构对节肢动物农业群落影响的认识,目前仅限于少数作物中少数物种的不同偏好识别(Kromp,1998;Honek和Jarosik,2000)。因此,在田间异质性如何影响节肢动物多样性及群落组成,以及多样性、丰度、体型分布等群落指标变化如何影响生物功能与生态系统服务供给方面,存在应用研究空白。
在有益节肢动物中,步甲被认为是重要的害虫天敌,包括谷物叶甲幼虫的捕食者(Kromp,1999;Meindl等,2001;Kheirodin等,2019)。谷物叶甲是欧、亚、北美主要谷物害虫,其幼虫可导致严重作物损害(Buntin等,2004),主要依赖杀虫剂防治。由于气候变化加剧叶甲与其寄生蜂间的物候失配,未来杀虫剂使用可能进一步增加(Evans等,2013)。此外,步甲是已知的重要杂草种子捕食者(Menalled,2006)。因此,能促进步甲且不带来农艺与经济劣势的作物管理措施,有望成为通过步甲提供生态系统服务实现冬小麦农药生态替代的范例。
本研究由此检验:简单调控作物结构(播种时保持部分开沟器关闭,不减少播种量且不影响常规耕作)是否会形成更异质的冬小麦田,从而对步甲生物多样性、害虫及杂草种子控制产生积极影响,且不降低产量。本研究旨在验证该措施除保护生物多样性外,是否具有生态集约化潜力。具体假设:1) 调控作物结构的田块比常规管理田块具有更异质的结构与小气候条件;2) 这与步甲群落更高多样性、活动密度及改变的体型分布相关;3) 可增强生态系统服务供给(害虫与杂草种子控制);4) 且不会导致显著产量损失。
2. 研究方法
2.1. 研究设计
为探究作物结构改变对步甲群落、害虫防治与种子捕食服务的影响,以及对禾谷类叶甲密度和作物损害的作用,本研究共设置了38块冬麦田(19块作物结构经过处理的麦田与19块未处理的对照麦田进行配对)。田间试验于2019年和2020年的4月至7月进行。19对采用常规管理的冬麦田(2019年6对,2020年13对)选自瑞士低地集约化农业景观区。每对麦田位置相邻,且具有相似的非生物与环境条件(如土壤类型、地下水位、气候条件及周边景观构成)。但无法保证每对麦田的小麦品种一致。作物管理措施(轮作前茬、土壤耕作、施肥、杀菌剂、除草剂与生长调节剂施用;未使用杀虫剂)基本一致,仅作物结构(即播种模式)存在差异。每对麦田中有一块作为对照田,播种时所有开沟器均保持开放,形成中欧常规小麦种植的均匀播种模式,行距因播种机型号略有差异(12.5–16.6 cm)(Abichou et al., 2019)。另一块麦田为作物结构处理组,通过关闭每4米工作宽度内的2–3个开沟器(具体数量取决于播种机型号及行距)改变播种模式,但不降低播种量(例如行距12.5 cm时:播种29行后留3行不播,以此类推;行距16.6 cm时:播种22行后留2行不播,以此类推)。因此,改造后的作物结构包含“未播种行”(开沟器关闭)和“密播行”(开沟器开放)。由于处理区播种量未变(400粒/平方米),原本应播于“未播种行”的种子被分配至“密播行”,导致后者播种密度略高(图1A)。麦田平均面积为4.93公顷(±3.36公顷),配对田块间最小距离为300米以确保独立性(平均值±标准误:733±482米),但有6对麦田因配对田块相似性优先而小于该距离。试验在三种样方类型(处理田的“未播种行”与“密播行”;对照田的“常规播种行”)及田块中心区域(避免边缘效应)进行。
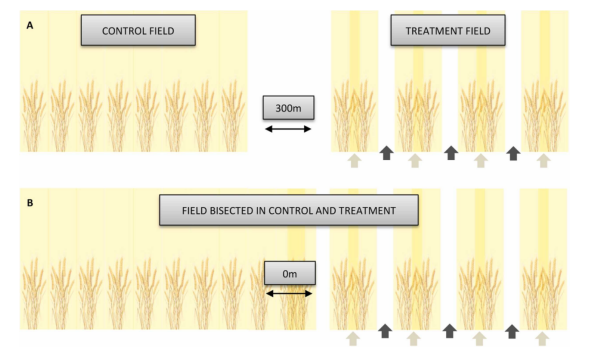
图1.冬小麦播种模式示意图(改造与未改造作物结构)
对照区(左)所有开沟器开放,形成“常规行”均匀播种模式。处理区(右)通过每4米工作宽度关闭2–3个开沟器(不减少播种量)改变播种模式,形成包含“未播种行”(开沟器关闭)和“密播行”(开沟器开放)的作物结构。图中深色箭头指示处理田的“未播种行”,浅色箭头指示“密播行”。对照区与处理区行距均为12.5–16.6 cm(因播种机而异),符合中欧常规小麦种植标准行距(Abichou et al., 2019)。(A) 独立对照田与处理田。为避免移动生物溢出效应,配对对照田与处理田最小间距为300米。所有涉及移动生物(步甲陷阱采样、捕食实验、害虫水平与作物损害观测)及相应微气候与结构测量的评估均在独立田块进行。(B) 分割田块(一半对照播种模式,一半处理播种模式)。为排除播种模式外其他因素对产量的影响,产量测量在同一田块内配对进行。
评估作物结构改变对产量影响时采用稍异设计。在17块不同麦田上测量两种作物结构类型下的产量:每块田一半区域设置处理组(改造结构),另一半为对照组(常规结构)(图1B),两半区域构成配对。这些田块于2017–2021年间在同一研究区域建立,管理措施同上,但每对田块内小麦品种相同(不同对间品种仍异)。田块平均面积为15.48公顷(±4.90公顷)。产量评估在两种样方类型(处理区的“未播种行”与“密播行”;对照区的“常规播种行”)中进行。
在同一田块内研究作物结构对产量影响的原因在于:减少不同田块间可能影响产量的变异因素(如养分水分有效性、小麦品种等)。而在不同田块研究作物结构对步甲及害虫防治与种子捕食服务的影响,并保持处理与对照田最小间距,是为了减少步甲等移动节肢动物的潜在交换及处理-对照田间的溢出效应。
2.2. 田间微气候与结构特征评估
为验证改造作物结构的田块是否呈现不同微气候与结构特征(假设1),本研究收集了温度、湿度与植被数据。
2020年在13对田块的三种样方类型中各安装温湿度记录仪(TMS-4, TOMST, 捷克),每15分钟同步测量土壤湿度(地表下6 cm)与地表温度(地表上2 cm)(Wild et al., 2019)。
2019–2020年4–7月每月评估19对田块中每块田的植被结构:每种样方类型设4个0.2×2.5 m子样方,目测小麦覆盖百分比。
2.3. 天敌采样
为验证作物结构改造是否会改变步甲群落的活动密度、物种丰富度及体型分布(假设2),使用陷阱(每田每种样方类型2个;漏斗直径10 cm;70%乙醇保存,陷阱间距≥10 m)在处理与对照田中采样。2019–2020年4月初至7月初进行4轮采样(每轮一周)。每轮每田每种样方类型分析一个陷阱。成虫鉴定至种级,依据文献记录体型大小和营养级(肉食性、杂食性或植食性)(详见表S1)。按营养级划分 guild:肉食-杂食guild用于害虫防治,植食-杂食guild用于杂草种子控制。
2.4. 替代猎物法测量天然害虫与杂草种子控制
2020年采用替代猎物(即哨兵)按QuESSA标准协议 测量处理与对照田捕食率。通过害虫捕食实验(昆虫哨兵)和杂草种子捕食实验(种子哨兵)收集数据,以分析作物结构改造对天然害虫与杂草种子控制的影响(假设3),并解析步甲群落指标(活动密度、物种丰富度、体型分布)与捕食功能间的关联机制。
测量处理与对照田捕食率。通过害虫捕食实验(昆虫哨兵)和杂草种子捕食实验(种子哨兵)收集数据,以分析作物结构改造对天然害虫与杂草种子控制的影响(假设3),并解析步甲群落指标(活动密度、物种丰富度、体型分布)与捕食功能间的关联机制。
2.4.1. 害虫捕食实验
选用蓝瓶蝇幼虫(双翅目)作为哨兵,因其符合广食性天敌捕食模式(自身观察),且比禾谷类叶甲幼虫更易操作(Kheirodin et al., 2020)。将10条三龄活幼虫用昆虫针(1号)固定于7.5×15 cm泡沫板制成“幼虫板”。每样方设两条3米样带,每条样带埋入3块与地表齐平的“幼虫板”,并用钉子固定。2020年6月末至7月初在13对处理与对照田中昼夜暴露12小时。共暴露9360条幼虫(13对田×3种样方×2次采样×3板/样带×10幼虫/板×2昼夜)。部分或全部取食记为被捕食。
同步在每条样带1米内设置陷阱采样,以评估活跃步甲捕食者群落。据此将活动密度、物种丰富度和体型分布与样带捕食率关联。鉴定方法同2.3节。仅分析体长≥5.0 mm的肉食性与杂食性步甲作为潜在捕食者。
2.4.2. 杂草种子捕食实验
选用荠菜(十字花科,0.1 mg/粒)、 Tripleurospermum inodorum(菊科,0.3 mg/粒)、繁缕(石竹科,0.4 mg/粒)、早熟禾(禾本科,0.4 mg/粒)和野芝麻(唇形科,0.8 mg/粒)五种重要农田杂草种子(Häni et al., 2008)。每物种20粒种子随机粘于4×9.5 cm砂纸(K80)制作的“种子卡”(Brust and House, 1988改进)。种子卡埋入土表下约5 mm并用钉子固定。2020年4–7月进行四轮暴露(每轮7天)。13对处理与对照田共设置60个样本单元(5物种×20粒×3样方类型×4重复),累计15600粒种子。回收后实验室双目镜下计数每物种被取食种子数。
“种子卡”放置与每周陷阱采样同步(距陷阱1米)。仅考虑体型2.5–8.0 mm的植食性与杂食性步甲作为潜在捕食者(因步甲取食种子大小受颚部尺寸限制)(Forsythe, 1982; Honek et al., 2007)。
2.5. 禾谷类叶甲密度与作物损害评估
除害虫捕食实验外,通过评估禾谷类叶甲密度及相关作物损害,可分析非实验条件下天然害虫控制水平(进一步支持假设3)。
禾谷类叶甲成虫取食幼嫩麦苗不影响产量,但幼虫啃食叶肉组织导致叶片 skeletonize,降低光合能力造成显著损害(Buntin et al., 2004)。
2020年在13对处理与对照田中,于幼虫发生高峰期(5月末/6月中)三次记录每种样方类型(“未播种行”样方选取邻近未播种行的小麦)2×25株小麦旗叶上的幼虫数量。但因当年幼虫发育提前,仅分析首次记录数据。6月中对同批小麦评估幼虫造成的旗叶损害百分比。成虫采样于7月早中期在13块(2020年)和4块(2019年)田中进行,采用标准扫网法(40网次,网径40 cm),以评估7月初由幼虫发育的第二代成虫密度,反映天敌对卵、幼虫和蛹的整体控制效果。
2.6. 作物产量评估
为收集不同作物结构下的产量数据(假设4),2017–2021年间使用联合收割机产量绘图功能对17块田进行产量调查(见2.1节)。基于土壤图、产量图和地形特征划分均质区域(Bobryk et al., 2016),按实验设计将相邻子样方归入对应作物结构类型。采用Quantum GIS 3.14 计算各子样方谷物产量标准化平均值(14.5%含水量)。
计算各子样方谷物产量标准化平均值(14.5%含水量)。
2.7. 统计分析
根据响应变量的误差分布,拟合了广义线性混合模型或线性混合模型。通过在每个模型中包含一个观测水平随机因子并将其与原始模型进行比较,来检查广义线性混合模型是否存在过度离散现象;若该因子能显著改善模型,则保留原始模型。高斯分布(正态分布)的模型假设根据 Zuur 等人 (2009) 推荐的图形验证程序进行检查。所有数值型解释变量在分析前均进行了标准化处理。所有模型均将嵌套在田对ID内的田间ID作为随机因子纳入。分析使用 R ver.4.0.2 (R Core Team, 2020) 进行。
对于具有生态学意义的变量及其效应(田间的微气候和结构特征、天敌、哨兵被捕食情况、谷物叶甲密度和作物损害),重点关注三种样方类型(对照田中的‘常规播种行’、处理田中的‘未播种行’、处理田中的‘密播行’),以分析样方类型的局部效应,并探究这三个区域如何在田间形成不同的生态位及其相应效应。分别使用‘常规播种行’与‘未播种行’、‘常规播种行’与‘密播行’之间的差异得出结论。对于产量,将整个处理区域(由‘未播种行’和‘密播行’组成)与对照区域(由常规播种行组成)进行比较,以获得每公顷产量的相关信息。
当 p < 0.05 时,效应定义为显著;当 0.05 ≤ p < 0.10 时,定义为边缘显著。
2.7.1. 田间的微气候和结构特征
为检验处理田是否提供了更异质化的结构和微气候条件(假设1),比较了来自TMS记录仪的数据和目测观察的不同样方类型的作物覆盖度。
使用土壤湿度和土壤表面温度作为单独的响应变量(假设其服从高斯分布),分析了具有和不具有 manipulated crop architecture (操控作物结构) 的田间的微气候条件。对温度进行了对数转换以获得正态分布的残差并避免异方差性。使用作物覆盖度作为响应变量(假设其服从高斯分布)分析了结构条件。模型将作物结构(三个水平:‘常规播种行’、‘未播种行’、‘密播行’)作为固定因子。数据收集的时间点嵌套在已建立的田对/田间ID随机因子中。
2.7.2. 天敌
为检验作物结构是否塑造了天敌群落,分析了活动密度、物种丰富度和群落加权平均体尺如何受作物结构的影响(假设2)。针对不同的捕食者群落分别对上述每个响应变量拟合了模型:肉食性-杂食性步甲(与害虫控制相关)、植食性-杂食性步甲(与杂草种子控制相关)。对活动密度和物种丰富度使用假定泊松分布的广义线性混合模型,对群落加权平均体尺使用假定高斯分布的线性混合模型。模型将作物结构(三个水平:‘常规播种行’、‘未播种行’、‘密播行’)作为固定因子。
2.7.3. 哨兵猎物的被捕食
分析了作物结构对哨兵被捕食的影响,以检验在具有 manipulated crop architecture (操控作物结构) 的田中,天然害虫和杂草种子控制是否得到增强(假设3)。分别将(被捕食或未被捕食的)蓝瓶蝇幼虫和杂草种子作为二元响应变量,假定数据服从二项分布。模型中包含了相关步甲群落的不同描述因子作为解释变量,以更好地理解它们在捕食事件中的作用。因此,蓝瓶蝇被捕食的模型将作物结构(三个水平:‘常规播种行’、‘未播种行’、‘密播行’)和昼夜时间(两个水平:白天、夜晚)作为固定因子,并将体尺至少为 5.0 mm 的肉食性和杂食性步甲的活动密度、物种丰富度和群落加权平均体尺作为连续变量。
杂草种子被捕食的模型将作物结构(三个水平:‘常规播种行’、‘未播种行’、‘密播行’)作为固定因子,并将体尺在 2.5 至 8.0 mm 之间的植食性和杂食性步甲的活动密度和物种丰富度作为连续变量。种子种类作为随机因子纳入。
物种丰富度、活动密度和群落加权平均值的数据分别来自每个‘幼虫板’和‘种子卡’相关联的陷阱样本。由于潜在杂草种子捕食者的体尺上限受到限制,群落加权平均体尺仅包含在害虫被捕食实验的分析中。
2.7.4. 谷物叶甲密度和作物损害
为检验在具有 manipulated crop architecture (操控作物结构) 的田中天然害虫水平是否降低(假设3),分析了作物结构对谷物叶甲幼虫、第二代谷物叶甲成虫和作物损害(各自作为单独的响应变量)的影响,假定误差服从泊松分布。模型将作物结构(三个水平:‘常规播种行’、‘未播种行’、‘密播行’)作为固定因子。
2.7.5. 作物产量
使用具有高斯误差分布的模型分析了作物结构对产量的影响(假设4),并将处理作为固定因子(两个水平:对照、处理)。
3. 结果
3.1. 作物结构对微气候和结构条件的影响
4月至7月期间,处理田‘未播种行’的土壤表面平均温度显著高于对照田的‘常规播种行’(平均高出5.9%),但这并未导致土壤湿度发生显著变化。同期,‘未播种行’的作物覆盖度显著低于‘常规播种行’(平均低92.2%)。‘密播行’和‘常规播种行’的微气候和结构条件彼此间无显著差异。模型摘要参见表1,原始数据的平均值±标准误参见表S2。
表1.解释不同作物结构(conventional 指对照田(无操控作物结构)中的‘常规播种行’;dense 指处理田(有操控作物结构)中的‘密播行’;unsown 指处理田中的‘未播种行’) 之间土壤表面温度、土壤湿度和作物覆盖度差异的混合线性模型摘要。Est. = 估计值, SE = 标准误, DF = 自由度。P ≤ 0.05 的值以粗体显示。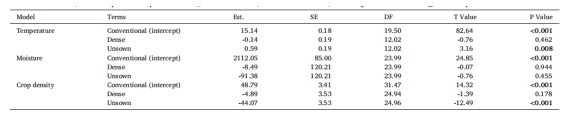
3.2. 作物结构对天敌的影响
在具有 manipulated crop architecture (操控作物结构) 的田中,潜在害虫捕食者的物种丰富度在‘未播种行’中比对照田的‘常规播种行’平均高出10.6%(边缘显著)(图2A),而活动密度在处理田和对照田的样方类型之间无显著差异。在‘未播种行’中捕获的潜在害虫捕食者群落的体尺平均比‘常规播种行’中大6.5%(显著;图2C)。模型摘要参见表2,原始数据的平均值±标准误参见表S3。
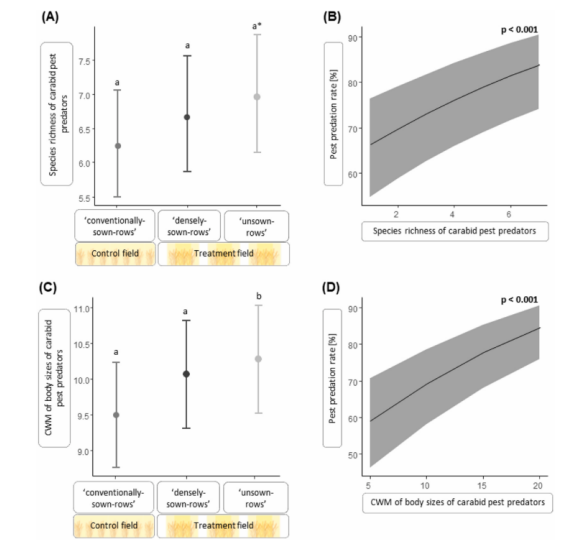
图2.不同冬小麦作物结构下步甲害虫捕食者群落的物种丰富度和体尺分布,以及这些群落描述因子如何影响害虫被捕食。(广义)线性混合效应模型的平均预测值。误差线显示89% CI。对照田中的‘常规播种行’具有均匀播种模式;处理田采用异质播种模式建立,包括‘未播种行’和‘密播行’。(A) 作物结构对步甲害虫捕食者物种丰富度的影响。‘未播种行’(a) 的物种丰富度边缘显著高于‘常规播种行’(a)。(B) 步甲害虫捕食者物种丰富度与被捕食的害虫哨兵百分比呈显著正相关。(C) 作物结构对步甲害虫捕食者体尺分布的影响。‘未播种行’(b) 的群落加权平均(CWM)体尺显著高于‘常规播种行’(a)。(D) 步甲害虫捕食者的体尺CWM与被捕食的害虫哨兵百分比呈显著正相关。*
表2.解释分别来自不同作物结构(conventional 指对照田(无操控作物结构)中的‘常规播种行’;dense 指处理田(有操控作物结构)中的‘密播行’;unsown 指处理田中的‘未播种行’) 的害虫捕食者(肉食性和杂食性步甲)或种子捕食者(植食性和杂食性步甲)的活动密度、物种丰富度和群落加权平均(CWM)体尺差异的(广义)混合线性模型摘要。数据来自每月(4月至7月)进行一次、持续一周的陷阱采样。Est. = 估计值, SE = 标准误, DF = 自由度。P ≤ 0.05 的值以粗体显示。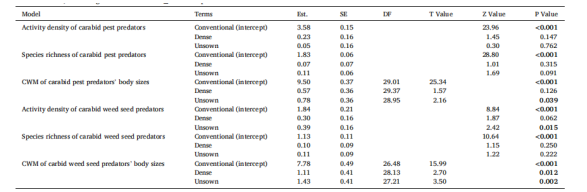
相反,与‘常规播种行’相比,潜在种子捕食者的活动密度在‘未播种行’(显著)和‘密播行’(边缘显著)中分别平均增加了45.5%和30.7%(图3A)。该功能团的物种丰富度未受作物结构的显著影响,但与‘常规播种行’相比,群落的体尺在‘未播种行’和‘密播行’中分别平均增加了13.9%和7.7%(均显著)。模型摘要参见表2,原始数据的平均值±标准误参见表S3。
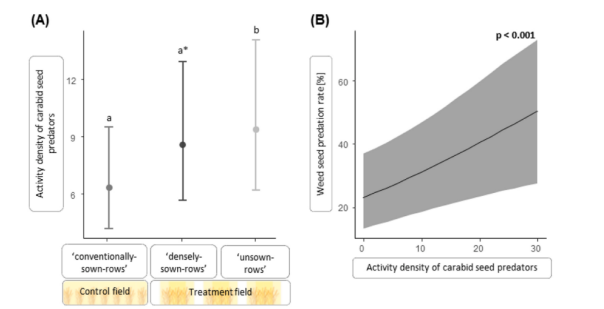
图3.不同冬小麦作物结构下步甲种子捕食者的活动密度及其与种子被捕食的关系。广义线性混合效应模型的平均预测值。误差线显示89% CI。(A) 作物结构对步甲种子捕食者活动密度的影响。对照田中的‘常规播种行’具有均匀播种模式;处理田采用异质播种模式建立,包括‘未播种行’和‘密播行’。活动密度在‘未播种行’或‘密播行’中显著(b)或边缘显著(a)高于‘常规播种行’(a)。(B) 步甲种子捕食者活动密度与被捕食的杂草种子哨兵百分比呈显著正相关。*
3.3. 作物结构及步甲群落对害虫和杂草种子捕食实验中自然控制的影响
与对照田的“常规播种行”相比,处理田的“密播行”中的害虫捕食率显著增加了平均10.1%(图4C)。夜间的捕食率平均比白天高5.0%(显著)。体型至少为5.0 mm的捕食性步甲更高的物种丰富度(图2B)、活动密度和体型大小(图2D)与哨兵害虫抑制率呈显著正相关。模型摘要参见表3,原始数据平均值±标准误参见表S4。
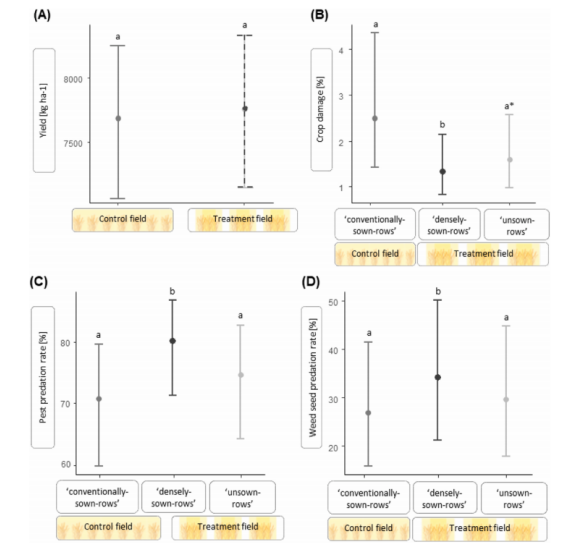
图4.冬小麦作物结构对产量、谷物叶甲造成的作物损害以及哨兵害虫和杂草种子捕食的影响。 图为广义线性混合效应模型的平均预测值。误差线显示89%置信区间(CI)。产量在对照田(均质播种模式,由“常规播种行”组成)和处理田(异质播种模式,由“未播种行”和“密播行”组成)中测量。作物损害以及害虫和杂草种子的自然控制则在对照田的“常规播种行”以及处理田的“密播行”和“未播种行”中测量。(A) 对照田和处理田的小麦产量无显著差异。(B) 以旗叶受损百分比衡量的作物损害,在“密播行”或“未播种行”中显著低于(b)或边缘显著低于(a*)“常规播种行”(a)。(C) 以被捕食的哨兵害虫百分比衡量的害虫捕食率,在“密播行”(b)中显著高于“常规播种行”(a)。(D) 以被捕食的哨兵杂草种子百分比衡量的杂草种子捕食率,在“密播行”(b)中显著高于“常规播种行”(a)。
表3.分别解释害虫和杂草种子哨兵捕食率差异的广义混合线性模型摘要。 分析了白天/夜晚、作物结构(conventional指对照田中‘常规播种行’(无操纵作物结构);dense指处理田中‘密播行’,unsown指处理田中‘未播种行’(有操纵作物结构))、物种丰富度、活动密度以及潜在害虫捕食者(体型至少5.0 mm的肉食性和杂食性步甲)的群落加权平均(CWM)体型大小对害虫哨兵(蓝蝇幼虫)捕食率的影响。分析了作物结构(conventional指对照田中‘常规播种行’(无操纵作物结构);dense指处理田中‘密播行’,unsown指处理田中‘未播种行’(有操纵作物结构))、潜在杂草种子捕食者(体型在2.5至8.0 mm之间的植食性和杂食性步甲)的物种丰富度和活动密度对杂草种子哨兵(荠菜(十字花科,每粒种子0.1 mg)、 Tripleurospermum inodorum(菊科,每粒种子0.3 mg)、繁缕(石竹科,每粒种子0.4 mg)、早熟禾(禾本科,每粒种子0.4 mg)、紫花野芝麻(唇形科,每粒种子0.8 mg))捕食率的影响。根据各自的实验设置,害虫捕食实验的步甲数据来自12小时陷阱样本,杂草种子捕食实验的步甲数据来自7天陷阱样本。Est. = 估计值, SE = 标准误。P ≤ 0.05 的值以粗体显示。
(表内容翻译略,保留英文术语如Est., SE, Z Value, P Value,并对行内注释进行翻译)
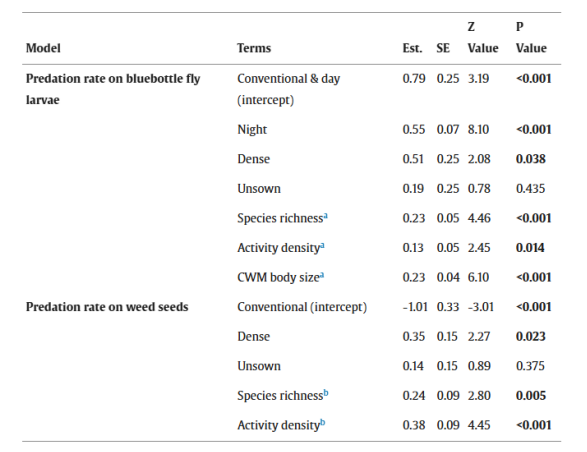
杂草种子捕食率在处理田中也显著高于对照田,同样仅在“密播行”中显著(平均提高6.5%;图4D)。体型在2.5至8.0 mm之间的潜在种子捕食者的物种丰富度和活动密度(图3B)对杂草种子抑制有显著正向影响。模型摘要参见表3,原始数据平均值±标准误参见表S4。
3.4. 作物结构对谷物叶甲密度和作物损害的影响
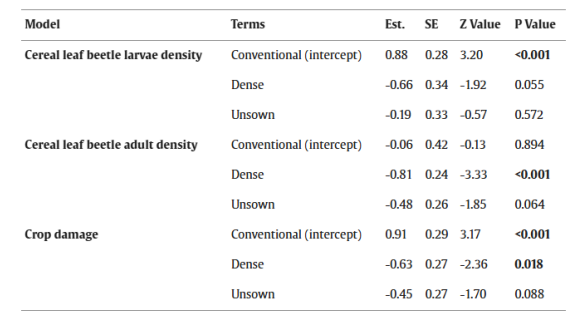
处理田“密播行”中的谷物叶甲幼虫数量与对照田的“常规播种行”相比,呈边缘显著减少(平均减少50%)。此外,与“常规播种行”相比,由谷物叶甲幼虫引起的作物损害以及再次出现的谷物叶甲成虫数量在“密播行”中显著减少,在“未播种行”中边缘显著减少(作物损害:‘密播行’减少56.3%;‘未播种行’减少33%;图4B。谷物叶甲成虫:‘密播行’减少52.1%;‘未播种行’减少34.1%)。模型摘要参见表4,原始数据平均值±标准误参见表S5。
表4.解释不同作物结构(conventional指对照田中‘常规播种行’(无操纵作物结构);dense指处理田中‘密播行’,unsown指处理田中‘未播种行’(有操纵作物结构))之间旗叶上谷物叶甲幼虫密度、第二代成虫及作物损害差异的广义混合线性模型摘要。 Est. = 估计值, SE = 标准误。P ≤ 0.05 的值以粗体显示。
3.5. 作物结构对产量的影响
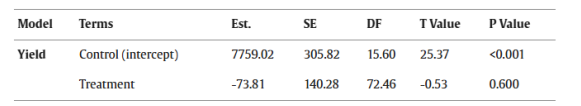
有无操纵作物结构的田块之间的产量无显著差异(图4A)。模型摘要参见表5,原始数据平均值±标准误参见表S6。
表5.解释无操纵作物结构的对照田(control)与有操纵作物结构的处理田(treatment)之间产量差异的混合线性模型摘要。 Est. = 估计值, SE = 标准误, DF = 自由度。P ≤ 0.05 的值以粗体显示。
4. 讨论
播种模式的改变导致了更异质的作物结构,这与步甲群落的变化、增强的害虫和杂草种子自然控制、降低的谷物叶甲密度以及减少的作物损害相关。同时,处理田的生产力得以维持。尽管本研究未直接测试不同农药投入水平,但这些结果表明了通过利用生物多样性来增强害虫和杂草种子自然控制服务以替代农药的潜力,尤其是在害虫密度高和作物损害严重的地区(Bommarco等人,2013)。然而,能在多大程度上替代,仍需进一步研究来验证。
陷阱取样和害虫捕食实验的结果表明,害虫控制效益(例如,减少的作物损害)可能是由捕食性步甲更高的物种丰富度和更大的体型所驱动的。这与先前的研究一致,表明农业害虫抑制可以通过大型步甲的存在(Rouabah等人,2014)和更高的天敌物种丰富度(Cardinale等人,2003)得到加强。根据Rouabah等人(2014)的研究,较大的步甲由于更高的人均消耗率可以增加害虫抑制。捕食者群落中已知的取样效应(Ives等人,2005)以及互补效应(参见Dainese等人,2019)可能分别是害虫抑制与物种丰富度呈正相关的原因。实际上,更多样化的捕食者群落可能增加的互补性(即,在目标食物项(即谷物叶甲卵、幼虫、蛹)方面的多样性增加)可能解释了为什么第二代谷物叶甲成虫的数量和作物损害分别比捕食实验中捕食者多样性与捕食率之间的负相关关系所预期的减少得更强。作物损害是谷物叶甲幼虫在田间发育期间取食小麦植株的结果,因此反映了天敌对谷物叶甲卵和幼虫的抑制。第二代谷物叶甲成虫还包含了蛹期作为捕食者的潜在食物项。另一方面,在害虫捕食实验中仅考察了对幼虫阶段的抑制。总而言之,结果表明操纵作物结构导致了更高效的害虫捕食者群落和增强的自然害虫控制服务,这可能有助于减少杀虫剂投入,从而降低农业生产的环境成本,但这需要在未来的研究中进一步验证。
杂草种子捕食者的活动密度对操纵的作物结构表现出强烈的正向响应,并与哨兵实验中增加的杂草种子捕食相关。这支持了先前一项研究的结果,该研究表明种子移除率与步甲种子捕食者数量呈正相关(Menalled等人,2007)。虽然活动密度在“未播种行”中最高,但种子捕食在“密播行”中最高。可能步甲利用“未播种行”进行无障碍移动,这增加了它们被陷阱捕获的可能性,而“密播行”则在取食活动期间提供了保护。总之,结果表明在具有操纵作物结构的冬麦田中,更高数量或更高活性的种子捕食者可能有助于减少除草剂的使用量。
操纵作物结构对步甲群落影响的一个潜在解释是,它们是由不同的作物密度引起的不同冠层内小气候条件所导致的。已知步甲受到土壤表面温湿度条件以及通过阳光照射进行体温调节能力的影响(Honek, 1988)。本研究的数据表明,处理田的“密播行”与对照田的“常规播种行”具有相似的 abiotic 条件。通常,稠密的作物株丛具有凉爽湿润的小气候,冠层下没有阳光斑(Honek and Jarosik, 2000)。相反,正如预期的那样,处理田的“未播种行”显著更温暖,阳光可到达土壤表面(本人观察),表明具有操纵作物结构的田块提供了更多样化的小气候条件。因此,这些异质条件可能导致了本研究中观察到的步甲群落变化,因为已知步甲对非生物条件具有物种特异性的偏好(Honek and Jarosik, 2000)。随着气候变化持续带来极端天气条件,能够提供不同小气候(微)生境的田块在未来可能变得更加重要。
为了通过生态集约化实际减少(而非仅仅转移)农业生产的环境成本,相关措施绝不能降低产量。先前的研究表明,小麦植株可以通过改变分蘖数或最终叶数等方式来补偿行距差异(Abichou等人,2019)。这可能解释了为什么在具有操纵作物结构的田块中没有产量损失:“密播行”中的单株小麦可能由于光照和水分获取减少而产量较低,但邻近“未播种行”的单株小麦可能由于资源获取增加而产量较高,预计两者可以相互抵消。然而,由于本研究范围不包括测试不同的农药施用方案,因此无法完全区分小麦植株的这种可塑性或更高的生物控制及相关的较低作物损害在维持产量方面的贡献程度。尽管如此,研究田块的作物损害和害虫水平从一开始就相对较低,因此在害虫水平较高的年份和地区,可以预期增强的自然控制会带来更强的积极效果。
从实践的角度来看,操纵作物结构对常规农事操作影响很小。播种前关闭播种开沟器是需要进行的(一次性)操作,使用标准播种机即可完成。无需对常规耕作方法进行进一步调整,也没有可能阻碍农民实施该措施的土地机会成本(Bailey等人,2015;Kleijn等人,2019)。因此,我们的研究结果应有助于并鼓励采用该措施,这也可能普遍提高对生态集约化措施的认识。
此外,可以预期操纵作物结构也可能成为促进其他服务(例如,授粉)以及用于其他表现出与小麦类似植物生长可塑性的作物(例如,向日葵和大豆(Junior等人,2018;Pereira and Hall, 2019))的一种有前景的方法。同样,其他受威胁物种,如地面繁殖的农田鸟类,也可能从该措施中受益,因为它在整个生长季在谷类作物田中创造了开放和可进入的区域(Wilson等人,2005)。此外,操纵作物结构与补充措施(如保护性农业(Palm等人,2014))相结合,可能会进一步促进生物多样性及相关生态系统服务。然而,需要进一步的研究来检验这些想法。
总之,所研究的作物结构操纵有潜力成为冬小麦生态集约化的有效工具,因为它显示了促进生物多样性和增强生态系统服务的效益,且不会带来农艺或经济上的劣势。特别是在害虫丰度高的研究区域,增强的自然害虫控制服务可能有助于农民减少农药施用,这将有助于降低单位产量的经济和环境成本,并可能帮助农民和农业部门实现农药减量计划,例如当前欧盟绿色协议所设定的目标(欧盟委员会,2019)。需要进行不同农药投入水平的进一步田间研究来检验这一点,并量化所研究的作物结构操纵作为小麦生产生态替代措施的潜力。