


精准太阳能跟踪系统对农田土壤生物多样性的影响
发布时间:2025-09-10 12:24 | 点击次数:135
精准太阳能跟踪系统对农田土壤生物多样性的影响
本文由上海赛弗生物科技有限公司翻译,旨在为相关科研人员提供科学参考,由于我司水平有限,错漏之处在所难免,敬请谅解指正!
原文题目:Impacts of punctual solar trackers on soil biodiversity in agricultural lands

亮点
• 草地与麦田中土壤生物多样性的响应略有差异。
• 土壤生物多样性受耕作实践的影响大于精准AV跟踪系统的影响。
• AV跟踪系统附近区域提供了未受干扰的土壤和植被条件。
摘要
可再生能源技术快速发展,太阳能是最具前景的能源之一。农业光伏能在同一块土地上实现作物种植与能源生产的结合。尽管许多研究探讨了地面安装太阳能电池板对未耕种草地的影响,但很少有研究关注农业光伏结构,尤其是采用二维转向支架且地面锚固有限的双轴跟踪系统。我们研究了此类跟踪系统(通过锚固对耕作实践的限制以及移动遮荫)对麦田和草地土壤物理、化学和生物特性的相对影响,并与已知影响这些特性的耕作实践进行了对比。通过PLS-PM分析发现,尽管跟踪器附近化学条件有所改变,且光伏结构带来了更高的特定植物丰富度,但与农业实践相比,其对生物群落并无显著影响。对比干草草地和麦田的结果表明,影响存在差异,需在不同农业背景下开展进一步比较研究。
关键词:光伏,农业光伏,跟踪系统,生物多样性,茶包指数(TBI,Tea Bag Index)
Photovoltaic,Agrivoltaic,Trackers,Biodiversity,TBI(Tea Bag Index)
1. 引言
未来几十年,可再生能源技术的发展预计将持续增长,以应对能源需求和减少温室气体排放的双重挑战(Dincer, 2000; Panwar et al., 2011)。其中,太阳能利用是最具前景的选择(IPCC, 2011),但其需要大面积的场地。尽管优先在屋顶、城市区域、棕地和人造基础设施上安装光伏(PV)板,这些场地仍十分有限。在此背景下,农业光伏(Agrivoltaics)作为一种有前景的方案应运而生,它能在同一块土地上实现农业种植与光伏发电的结合(Dupraz et al., 2011)。此外,它还可为农业提供多种服务,例如保护作物免受气候变化和气象灾害的影响、增加农场收入并提升能源自给能力。尽管农业光伏被认为可与农业协同发展,但它也引发了其他环境和社会问题,需谨慎应对以确保可持续发展(Hernandez et al., 2019)。
在此框架下,现有研究多集中于地面安装的太阳能电池板。例如,研究表明,由于太阳能电池板下的太阳辐射减少,白天的土壤温度降低(Armstrong et al., 2016; Marrou et al., 2013; Weselek et al., 2021),表明其在日尺度上具有缓冲效应(Barron-Gafford et al., 2019)。电池板下的土壤湿度较高,这很可能与温度降低导致的蒸散减少有关(Marrou et al., 2013),也可能与风速降低有关(Armstrong et al., 2016)。这些微气候效应可能对作物产量产生正面或负面影响,具体取决于环境条件(Weselek et al., 2021)。
在草地生态系统中也报道了类似结果:一些研究中,遮荫效应有助于维持较高的土壤湿度水平,从而提高了植物生物量(Hassanpour Adeh et al., 2018);而另一些研究中,微气候变化导致生物量减少(Armstrong et al., 2016; Lambert et al., 2023)。与光伏装置的农艺影响相比,其对生物多样性的影响迄今很少受到关注(Menta et al., 2023; Schweiger and Pataczek, 2023; Uldrijan et al., 2022),尽管生物多样性在多种农业生态过程中扮演重要角色(例如土壤有机质和碳储存、养分循环和作物害虫调控(Bardgett and Van Der Putten, 2014; Jouquet et al., 2014))。现有研究多涉及草地。一些研究表明,太阳能电池板的存在降低了板下的植物物种丰富度(Bai et al., 2022; Lambert et al., 2023)、土壤微生物活性(Lambert et al., 2021; Moscatelli et al., 2022)或生物量(Lambert et al., 2023),以及土壤节肢动物的丰度(Lambert et al., 2023; Menta et al., 2023)。由于土壤生物对微气候条件变化的敏感性(Barnett and Facey, 2016),这些研究得出结论:光伏电池板下降水模式和空气循环的改变引发了一系列过程,导致土壤肥力(包括有机质水平)的空间异质性分布不均(Moscatelli et al., 2022)。光伏装置对微气候的影响程度,进而对上述效应的影响,可能取决于光伏技术,例如其移动性/跟踪能力(Suuronen et al., 2017)和电池板高度,这些因素会改变阴影大小及其空间动态以及空气循环。因此,本研究涉及的跟踪系统对微气候的影响预计低于固定式太阳能电池板。相比之下,光伏装置在一年生农田中对生物多样性的影响仍然 largely unknown,尽管约50%的光伏基础设施预计将是在农业用地上安装的地面系统(Chatzipanagi et al., 2023; Dinesh and Pearce, 2016)。
在农田中,农业实践强烈影响土壤生物多样性,既通过直接减少生物活力甚至杀死生物(例如使用农药或耕作),也通过改变土壤物理和化学性质间接产生影响(Campbell et al., 2017; Hussain et al., 2009; Lichtenberg et al., 2017; Tsiafouli et al., 2015)。化学性质(如土壤有机碳和氮含量、pH值)和物理性质(如土壤压实度和湿度、矿物颗粒大小)直接影响土壤肥力和植物生长(Bronick and al, 2005),进而可能影响其他类群(如蚯蚓(Edwards and Arancon, 2022)、节肢动物或微生物(Coleman et al., 2017))的丰度和功能关系(Menta and Remelli, 2020)。
本研究旨在评估精准太阳能跟踪系统对农田土壤生物多样性的直接和间接(通过土壤及其植被覆盖的物理和化学特性)影响,同时考虑农业实践,特别关注遮荫和地面锚固的影响。具体而言,我们假设跟踪系统附近由于遮荫引起的微气候变化、自发植被的发展和安装过程中的土壤扰动,物理和化学条件将发生改变,从而影响土壤群落的活动。以法国西部的干草草地和冬麦田为研究对象,我们探讨了太阳能跟踪系统如何影响环境条件,并随后使用偏最小二乘路径模型(PLS-PM)量化了跟踪系统和农业实践对土壤生物多样性的直接和间接影响的相对重要性。
2. 材料与方法
11个研究点位于法国西部的布列塔尼和卢瓦尔河地区,这些地方的太阳能跟踪系统已安装二至七年:五个点在干草草地,六个点在冬麦田。调查于2023年3月14日至8月21日进行,期间平均气温、日降雨量和太阳辐射分别为16.02°C、3.02 mm和219Wm⁻²(气象数据:Meteomatics Weather API)。实验中心(雷恩)在此期间的平均云量为4.5 oktas.土壤类型在布列塔尼主要为棕壤(包括砂岩、页岩和风成粉砂),在卢瓦尔河地区主要为钙质土.
我们研究了Groupe OKwind®的光伏跟踪系统(图1A),其由117平方米的光伏板组成,支撑在7米高的桅杆顶部,因此下方仍可进行种植。光伏板通过双旋转轴在白天移动以跟踪并始终面向太阳。面板由双面单晶太阳能电池组成。这些跟踪系统的占地面积较小(6.25平方米),但混凝土基础附近无法耕作,使得杂草得以生长,尤其是在农田中(图1B)。

图1. (A) 跟踪系统的技术规格;(B) 混凝土基础附近难以耕作对植被的影响。
2.1. 野外调查与生物多样性数据收集
除两个干草草地因农业作业干扰无法评估微生物活性外(参见补充材料S1和S2),其余11个站点均采用了统一调查方案。每个站点选取一个太阳能跟踪系统进行研究。当存在多个跟踪系统时,优先选择外部干扰最小(如树篱或建筑物遮荫、相邻田地边缘效应、地面变形等)的个体。我们在桅杆基础三个不同距离和方位进行采样(图2):
(i) 北向与南向1米处(始终处于遮荫区,麦田中该区域未耕作且杂草丛生);
(ii) 北向8米处(每日仅部分时间遮荫);
(iii) 南向8米处(始终无遮荫);
(iv) 距边缘与跟踪系统同等距离的35米处(对照点,位于跟踪系统影响区外,不受遮荫(图3)及安装作业影响);
共设置四个采样位点。所有边缘与跟踪系统及对照点的最小距离均为20米。
2.1.1 蚯蚓丰度
通过人工分拣法采集蚯蚓样本:在每个采样点选取一个土壤块(长×宽×高:20×20×25厘米),在3月14日至4月6日期间分两个不同时间段进行采样,两次采样至少间隔两周。根据Bouché(1972)的分类方法,将蚯蚓划分为四个生态类群:表栖型、红色深栖型、黑色深栖型和内栖型。统计分析时对两个采样期数据分别处理。
2.1.2 节肢动物活动密度
使用直径9厘米的陷阱式捕捉器收集地表活动的跳虫、潮虫、马陆(Julida目)和球马陆(Glomerida目)。每个采样点设置两个相距4.5米的捕捉点,陷阱内注入200毫升盐水(饱和盐溶液)并加入数滴洗涤剂以降低表面张力,顶部加盖铝制防雨棚防止枯落物和雨水进入。捕捉器连续放置7天后收集,分三个时段进行:
(i)4月17日至4月28日(除一个干草草地外的所有站点);
(ii)5月2日至5月24日;
(iii)6月12日至6月27日。
个体鉴定到目级水平(Geoffroy, 1993; Hopkin, 2007; Sutton, 2013),并量化各类群的活动密度。统计分析时分别处理两个陷阱(采样点)和三个采样期的数据。
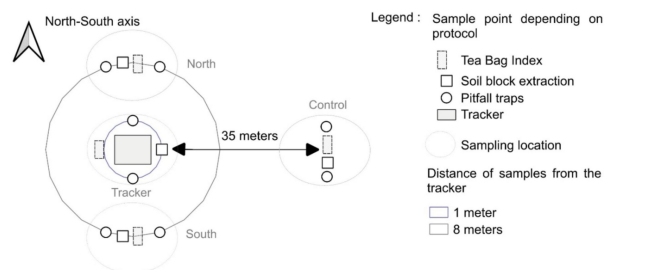
图2. 采样设计说明 每个采样点包含:用于评估微生物活性的绿茶包(green tea bags)与路易波士茶包(rooibos tea bags);用于测量蚯蚓丰度的土壤块取样;用于测量节肢动物活动密度的两个陷阱式捕捉器。
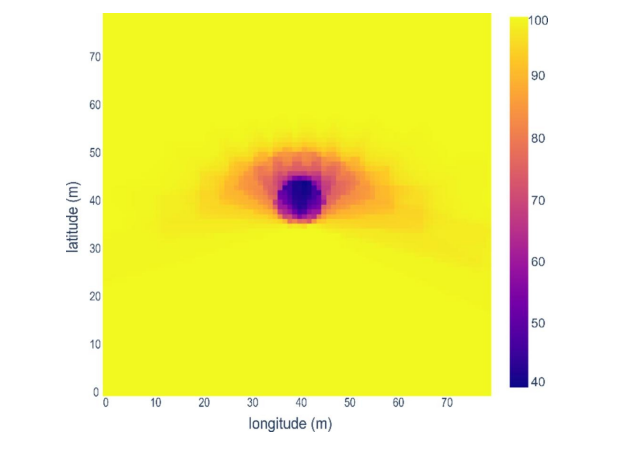
图3. 采样期间跟踪系统附近地面接收总辐射比的空间模拟(%RR色标)
本模拟案例基于雷恩地区(研究区域中心的最大城市及气象站所在地)219W/m²的太阳辐射数据。桅杆对应中心像素点。图示表明:从35米外开始,遮荫效应完全消失。
2.1.3. 土壤微生物活性
为测量有机质的分解速率,我们采用了茶包指数(Tea Bag Index,TBI)方法,这是一种用作土壤微生物活性指标的标准方案(Keuskamp et al., 2013)。将三包立顿绿茶(green teas)(EAN 87 10,908 90,359 5)和三包立顿路易波士茶(rooibos tea)(EAN 87 22,700 18,843 8)称量其干燥洁净时的重量,然后在每个采样点(每个田地 n = 12,总计)埋入土壤 8 厘米深处,放置 45 ± 3 天(总计从 6 月 5 日到 8 月 22 日)。之后取出茶包(Tea Bag),清理干净,并在 75 °C 下干燥 48 小时或直至茶包重量恒定。所有麦田采样点总共埋设了 60 个绿茶包(green tea bags)和 60 个路易波士茶包(rooibos tea bags),而所有草甸采样点则埋设了 36 个绿茶包和 36 个路易波士茶包。我们计算了两个 TBI 参数:稳定化程度(S),即凋落物分解的程度,反映了培养后剩余的未分解、可水解的不稳定部分的比例;以及分解速率(k),即植物材料的不稳定部分被分解的程度和速率(Keuskamp et al., 2013)。稳定化导致实际分解部分 a 偏离可水解部分 H,可以解释为环境条件对不稳定部分分解的抑制作用:
(1)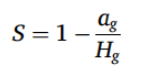
其中 ag 是绿茶(green tea)的可分解部分,Hg 是绿茶(green tea)的可水解部分。路易波士茶(rooibos tea)的可分解部分 (ar) 由其可水解部分 (Hr) 和稳定化因子 S 计算得出:
(2)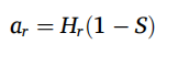
分解速率 k 使用以下指数衰减函数计算:
(3)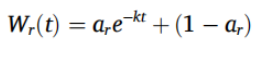
其中 Wr(t) 是培养时间 t 后路易波士茶(rooibos tea)的重量,ar 是不稳定部分,1 – ar 是路易波士茶的难分解部分。每个采样点单个茶包的 S 和 k 值在统计分析中分别保留。
2.2. 环境变量
2.2.1. 农业管理
通过对所有田地进行标准化农户调查,记录了 2022–2023 种植年度的农业实践。对于冬麦田,收集了以下数据:氮肥施用量 (kg.ha−1)、农药施用次数、耕作强度。对于干草草甸,收集了以下数据:刈割次数、刈割间隔平均时间、肥料类型(按强度等级排序:无肥 = 0;泻湖废水 = 1;粪肥 = 2;矿物氮肥 = 3)。
2.2.2. 土壤采样与理化分析
为表征土壤的物理和化学肥力,于 2023 年 3 月中下旬期间,在每个采样点使用深度为 20 厘米的土钻采集了 500 克新鲜土壤样品。样品经干燥后,由法国 AIA 阿拉斯土壤分析实验室进行分析。对粗颗粒部分(砾石、卵石)和细颗粒部分(粉砂、沙和粘土的比例)进行了定量 (g.kg−1)。从细颗粒部分测量了总石灰石 (CaCO3)、有机碳 (OC) 和总氮 (Ntot) 的含量 (g.kg−1)。在田间,我们还使用紧实度计在 8、16、24 和 32 厘米深度测量了土壤紧实度 (kPa; Spectrum Technologies, Inc.),使用 pH 计在土壤表面测量了土壤 pH 值 (Extech Instruments),并在每次访问采样点时使用 TMS-4 数据记录仪 (TOMST) 测量了土壤湿度。对于土壤湿度,基于三次记录计算 45 分钟的平均值。将原始湿度信号(50–200 MHz)进行转换并缩放到 1–4095 的数值范围(原始 TDT 数据)。为了将此相对值转换为体积土壤湿度 (%),采用了基于 (Wild et al., 2019) 的特定校准方法。
2.2.3. 植被
在冬麦田和干草草甸中,于三月、四月、五月和六月在每个采样点测量了 1 平方米区域(采样点两侧各 0.5 米)的植物高度。植物物种丰富度是基于 2023 年 5 月至 6 月期间一次对以两个 1 平方米样方为中心的 8 平方米区域内所有植物的识别来计算的。仅在冬麦田中,于三月、四月、五月和六月量化了采样点两侧各 0.5 米、共 1 平方米区域的植被覆盖度。
2.2.4. 遮荫条件
我们估算了每个采样期内每个采样点的遮荫强度,计算为给定时期内到达地面的累积辐射与同一时期无追踪器遮荫情况下理论接收累积辐射之比的倒数 (%RR),方法同 (Noirot-Cosson et al., 2022),使用了每小时辐射数据 (Meteomatics Weather API) (图3)。
2.2.5. 小气候条件
我们实施了一项平行实验,利用遮荫强度与相对湿度以及温度的关系,将土壤和空气温度以及相对空气湿度数据与我们测量的变量联系起来。在该实验中,于 2023 年 3 月 18 日至 7 月 7 日期间,在布列塔尼的四个冬麦田中,使用 TMS-4 数据记录仪 (TOMST) 记录了局部空气(地上 15 厘米)和土壤(地下 5 厘米)的每小时温度。数据记录仪均匀分布在四个追踪器周围的八个主要方位上,距离桅杆 5、10、15、20 和 35 米,以覆盖所有遮荫条件。我们在田间采样活动的同时记录这些微气候数据,即每月十五天:从 3 月 18 日到 4 月 1 日记录蚯蚓数据;从 4 月 15 日到 4 月 30 日,5 月 5 日到 5 月 20 日,6 月 1 日到 6 月 15 日记录节肢动物数据;从 6 月 15 日到 6 月 30 日记录微生物数据。我们通过平均两个最近样地的测量数据,并将追踪器南侧 35 米处的记录用作对照样地,来推断我们四个采样样地的小气候数据。
2.3. 统计分析
所有统计分析均在 R 版本 4.3.3 (R. Core Team, 2024) 中进行。冬麦田和干草草甸的数据分开分析。
2.3.1. 采样点的影响
2.3.1.1. 环境与生物学分析
使用线性混合效应模型 (LMMs) 或广义线性混合效应模型 (GLMMs) 以及 glmmTMB 包 (Magnusson et al., 2017) 量化了采样点 (n = 4) 对环境变量和土壤生物多样性的影响。对所有环境变量使用高斯误差分布模型,但土壤湿度除外(使用 Beta 分布模型),植被覆盖度、石块和砾石也除外(使用 Tweedie 分布模型)。对所有土壤生物多样性变量使用泊松分布模型,但稳定化程度 (S) 除外(使用高斯误差分布模型),分解速率 (k) 也除外(使用 Beta 分布模型)。使用 fitdistrplus 包 (Delignette-Muller and Dutang, 2015) 的函数 ‘fitdist’ 和 DHARMa 包 (Hartig, 2017) 选择分布族。我们在所有模型中纳入田地 ID 作为随机截距项,以解释同一田地里采样点之间潜在的空间自相关性。当结果显著时(p 值 < 0.05),使用 emmeans 包 (Lenth, 2020) 进行 Tukey 多重比较检验,以检测哪些采样点彼此之间存在差异。
2.3.1.2. 小气候分析
使用与前一节相同的程序,通过线性混合效应模型 (LMMs) 量化了两个效应。首先,遮荫强度对小气候条件的影响;其次,采样点对小气候条件的影响。两个模型均使用高斯误差分布模型。我们在所有模型中纳入田地 ID 作为随机截距项,以解释同一田地里采样点之间潜在的空间自相关性。当结果显著时(p 值 < 0.001),应用 Tuckey 多重比较检验来检测哪些采样点彼此之间存在差异。
2.3.2. 太阳能追踪器与农业管理对土壤生物多样性的各自重要性
2.3.2.1. 环境变量间的关系
使用 Pearson 检验和 corrplot 包 (Wei et al., 2017) 量化了环境变量之间的相关性,并分别对麦田和草甸使用 factoextra 包 (Kassambara, 2017) 通过主成分分析 (PCA) 进行图示。为避免高度相关变量在 PCA 中过度代表,我们移除了相关性高于 0.80 阈值的变量。这些结果也用作 PLS-PM 分析(见下文)中定义潜变量的初步方法。
2.3.2.2. PLS-PM 分析
使用偏最小二乘路径模型 (PLS-PM) 分析 (Sanchez, 2013) 对 (i) 农业实践、(ii) 追踪器对遮荫强度的影响、(iii) 追踪器基部附近的环境条件、(iv) 植被、(v) 土壤物理性质、(vi) 土壤化学性质以及 (vii) 土壤生物多样性之间的关系进行了建模。PLS-PM 是路径分析的一部分,路径分析是检验和测量变量组之间多重联系和影响的多元方法 (Tenenhaus and Tenenhaus, 2011)。这种方法特别适用于小样本 (Chaput, 2007)。
PLS-PM 由两个子模型组成,即内部模型和外部模型。外部模型描述了一组显变量 (MVs)(即可观测变量)与由这些显变量构建的合成潜变量 (LV) 之间的关系(通常代表无法直接测量的概念)。LV 及其相关的 MVs 形成一个称为“块”的组。接着,内部模型描述了 LV 之间的关系。在我们的研究中,PLS-PM 由七个块组成:
“农业管理” LV:对于冬麦,由以下 MVs 构建:氮肥施用量、农药施用次数、耕作强度;对于干草草甸,由以下 MVs 构建:刈割次数、刈割间隔平均时间、肥料类型。
由于追踪器对环境有不同类型的影响,我们将相关变量分成两个 LVs。因此,“遮荫条件” LV 由以下 MV 构建:遮荫强度;“追踪器基部区域” LV 基于一个二元 MV 构建,该 MV 定义为代表与追踪器结构的接近程度(0 = 大于 1 米;1 = 距离追踪器 1 米)。由于 PLS-PM 模型基于普通回归,它们可以处理二元变量 (Jakobowicz, 2008)。
“植被” LV:对于麦田,由以下 MVs 构建:植被覆盖度、植物物种丰富度、植物高度;对于干草草甸,由以下 MVs 构建:平均高度和物种丰富度。
“土壤物理性质” LV:由以下 MVs 构建:8、16、24 和 32 厘米处的土壤紧实度、石块、砾石、粘土、粉砂和沙含量,以及土壤湿度。
“土壤化学性质” LV:由以下 MVs 构建:总碳、氮、有机质和石灰石含量、C/N 比和 pH 值。
“土壤生物多样性” LV:通过更改“土壤生物多样性”块的内容,为麦田和草甸分别构建了三个独立的模型。对于蚯蚓丰度,该块由表栖型、内栖型、黑色孔栖型和红色孔栖型蚯蚓的个体丰度表示。对于微生物活性,该块由分解速率 (k) 和稳定化程度 (S) 指标表示。对于节肢动物活动密度,该块由跳虫、潮虫和马陆的个体活动密度表示。
我们模拟了除农业管理、遮荫条件和追踪器基底面积之外的所有潜变量(LVs)之间的关系。由于我们预期某些相关的潜变量之间存在强关系,因此我们进行了以下关联:
农业管理与植被、土壤物理性质和土壤化学性质相关联,因为农业用地不可避免地会导致土壤特性的变化,如养分状况、pH值、有机质含量、杂草多样性以及物理性质(Powlson等,2011)。
遮荫条件与上述相同的模块相关联,因为它影响微气候条件,包括温度、空气湿度、土壤湿度和光照条件(Armstrong等,2016;Tanner等,2020),因此可能影响光伏板下的土壤过程和植物群落(Lambert等,2021)。
追踪器基底面积与上述相同的模块相关联。混凝土基座底部的区域受到太阳能追踪器安装的严重影响,无法耕作,因为这会破坏结构以及农业设备。这里可能会形成特定的生态和环境条件,导致特殊的土壤状况和杂草发展。
土壤生物多样性潜变量与其他六个潜变量相关联。
鉴于研究目标并避免潜变量之间过多的关联,我们没有在植被、土壤物理性质和土壤化学性质之间建立关联。使用相关系数来衡量两个潜变量之间的关联,每个测量变量(MV)与其相关潜变量之间的关系被视为线性回归。土壤生物多样性模块以形成性方式构建,意味着潜变量由其测量变量构成。所有其他模块以反映性方式构建,意味着测量变量由其潜变量引起或反映。
最终的偏最小二乘路径模型(PLS-PM)通过以下三个步骤构建(Sanchez,2013):
初始模型拟合了一个反映田间功能和所有可能测量变量的固定内部模型(图4A);
在第二个模型中,我们确保了潜变量的单维性(即与一个潜变量相关的所有测量变量必须与其正相关)。然后,所有与潜变量负相关的测量变量乘以-1,以在运行最终模型之前保持单维性(图4B);
在最终模型中,与潜变量弱相关(即<0.4;Sanchez,2013)的测量变量被移除(图4C)。模型质量通过两个指标评估:拟合优度(GoF)作为内部和外部模型之间的折衷(每个模块的平均共同性×平均R²),若>0.70则认为良好;以及依赖潜变量的R²,若≥0.5则认为中等良好。为了确认模型在变量共线性方面的有效性,我们计算了方差膨胀因子(VIF)并验证每个模型的VIF<10(Bennaceur和Chafik,2019;Lacroux,2009)。
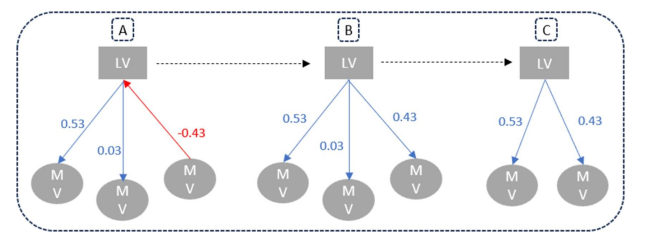
图4. PLS-PM分析的三步流程
3. 结果
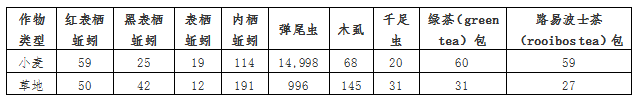
共收集了549条蚯蚓、15,994只弹尾虫、213只木虱和51只千足虫。在排除损坏的茶包后,回收了91个绿茶包和86个路易波士茶包(表1)。
表1. 每个分类群的样本个体总数或绿茶(green tea)/路易波士茶(rooibos tea)包的数量
3.1 采样位置对生物多样性和环境条件的影响
3.1.1 环境和生物变量
大多数生物多样性和环境变量受采样位置的影响,而与土地覆盖类型无关。在冬小麦田和干草草地中,植物物种丰富度和遮荫强度在距离追踪器1米处较高。相反,pH值、土壤氮、碳和有机质含量在距离追踪器35米处较高(在冬小麦田中,除pH值外,8米南也是如此)。在冬小麦田中,杂草覆盖率在1米处较高,而植物高度较低;土壤石灰含量在35米处最高。最后,在干草草地中,土壤C/N比在距离追踪器1米和8米北较低,而土壤石灰含量在1米处较高。更多详情见补充材料,表S3为统计结果,图S3.1至3.6为结果图表。
在生物多样性方面,蚯蚓丰度和千足虫活动密度在不同采样位置之间没有显著差异,而与土地覆盖类型无关,而木虱活动密度(图5)在小麦田和干草草地中均在距离追踪器1米处较高。在冬小麦田中,弹尾虫活动密度在1米处较低,而分解率(k)(图6)和稳定化(S)在35米处较低。在干草草地中,弹尾虫活动密度和分解率(k)在35米和8米北较高。更多详情见补充材料,表S3为统计结果,图S3.7为结果图表。
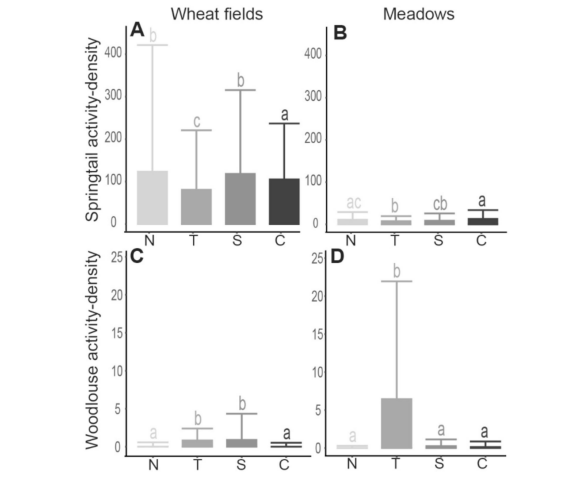
图5. 冬小麦田与干草草甸中跳虫(A-B)和潮虫(C-D)的平均活动密度(均值±标准差)。字母表示通过Tukey检验得出的不同位点间显著性差异(p < 0.05)。N:北向8米处;T:追踪器基座1米处;S:南向8米处;C:35米对照点。
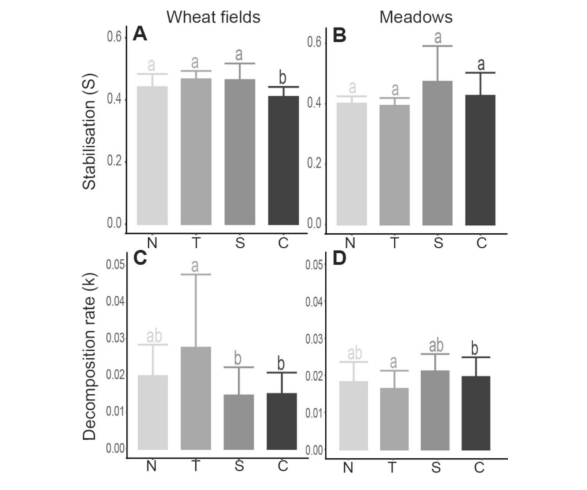
图6. 冬小麦田与干草草甸中稳定化指数(S;A-B)和分解速率(k;C-D)的平均值(均值±标准差)。字母表示通过Tukey检验得出的不同位点间显著性差异(p < 0.01)。N:北向8米处;T:追踪器基座1米处;S:南向8米处;C:35米对照点。
3.1.2 小气候
在模型1和模型2中,所有变量的结果均显著(表2)。模型1显示,在追踪器北侧1米和8米处的相对湿度差值较高(图7)。追踪器北侧1米和8米处的土壤温度与气温较低。与对照组相比,追踪器基座处的温湿度变化比北侧位置更为显著。
表2. 线性混合模型结果
微气候参数的位置效应与遮阴效应广义线性混合模型(GLMM)结果分析
|
参数 |
模型1 - 遮阴强度 |
模型2 - 位置 |
||
|
卡方值 |
P值 |
卡方值 |
采样点 |
|
|
遮阴强度 |
- |
- |
25,949 |
<0.001 |
|
相对湿度差 |
5820.8 |
<0.001 |
5820.8 |
<0.001 |
|
土壤温度差 |
286.28 |
<0.001 |
29,066 |
<0.001 |
|
气温差 |
586.27 |
<0.001 |
1664.3 |
<0.001 |
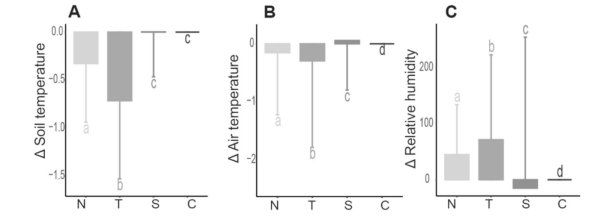
图7. 温度(℃;A-B)和相对湿度(原始TDT数据;C)的平均值。字母表示通过Tukey事后检验得出的不同位点间显著性差异(p < 0.01)。N=北侧,T=追踪器基座,S=南侧,C=对照组。
3.2. 太阳能跟踪器与农业管理对土壤功能的相对重要性
3.2.1. 环境变量之间的关系
在进行PCA分析之前,由于强相关性(r > 0.80),我们移除了冬小麦田23个变量中的5个(表S4.1)以及干草草甸22个变量中的6个(表S4.2)。在冬小麦田和干草草甸中,16厘米处的土壤紧实度被排除,因为该变量与8厘米和24厘米处的土壤紧实度相关;氮和碳含量也被排除,因为它们与有机质含量相关。仅在冬小麦田中,石块和砂粒含量被排除,因为它们与砾石含量相关。仅在干草草甸中,32厘米处的土壤紧实度被排除,因为该变量与24厘米处的土壤紧实度相关;石灰岩含量和平均割草间隔时间也被排除,因为它们分别与pH值和割草次数相关。
对于冬小麦田,我们保留了前两个PCA轴,它们解释了总方差的42.6%(图8A;表S5.1)。第一轴(24.5%)将具有高耕作强度、高植物物种丰富度和高碳氮比的采样点与具有高土壤紧实度、高粉粒和高氮含量的采样点区分开。第二轴(18.1%)主要将具有高土壤氮含量、高有机质含量、高土壤紧实度和高植物高度的采样点与具有高粉粒含量的采样点对立起来。跟踪器基座(tracker basis)的重心(barycentre)位置与其他位置明显不同。
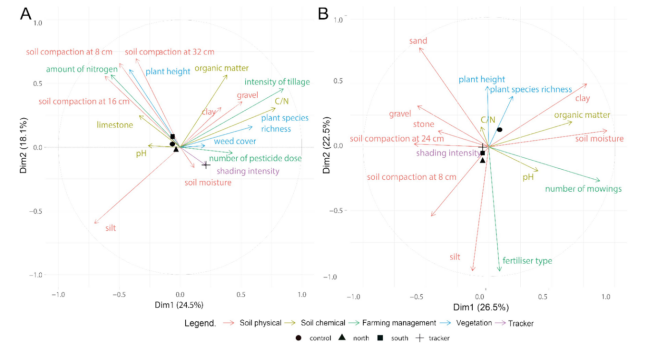
图8. 主成分分析(PCA)结果,展示了本研究中考虑的环境变量之间的关系,分别针对冬小麦田(A)和干草草甸(B)。颜色组对应于后续PLS-PM分析中考虑的潜变量(latent variables)。符号对应于每个采样点的重心。
在干草草甸中,我们也保留了前两个PCA轴,它们解释了总方差的49.0%(图8B;表S5.2)。轴1(26.5%)主要将割草次数最多、土壤最湿润、粘粒和有机质含量最高的采样点,与土壤更深处紧实、砂粒和砾石含量更高的采样点区分开。轴2(22.5%)主要将土壤表层更紧实、粉粒含量更高、进行有机施肥的采样点,与砂粒含量更高的采样点对立起来。对照组(control plots)的重心与其他位置完全分开。
3.2.2. 偏最小二乘路径模型(PLS-PM)
所有六个PLS-PM模型(分别针对冬小麦田或干草草甸中的蚯蚓丰度、节肢动物活动密度和微生物活性)的GoF系数至少为0.52(表S6),对应于良好的模型预测(Sanchez, 2013)。交叉载荷(cross-loadings)的结果表明,所有反映型区块(reflective blocks)在每个模型中都是有意义的且构建良好(表S7)。
3.2.2.1. 蚯蚓丰度
冬小麦田的PLS-PM模型(图9A)显示,蚯蚓丰度的大部分变异归因于红色表层种(red anecic)和内栖种(endogeic)蚯蚓。内部模型显示,蚯蚓丰度由其潜变量(LVs)很好地描述(R2 = 0.33),但没有LV与蚯蚓丰度显著相关。所有反映型LV都由其显变量(MVs)非常好地描述(ρ > 0.83)。植被由其相关的外生LV(exogenous LVs)非常好地解释(R2 = 0.81),主要受到位于跟踪器基面积(tracker basal area)内的正向影响,并受到农业管理的负向影响。土壤物理性质由其相关的外生LV非常好地解释(R2 = 0.83),主要受到农业管理的正向影响。土壤化学性质由其相关的外生LV较好地解释(R2 = 0.33),主要受到农业管理的负向影响。当同时考虑农业管理和跟踪器相关LV的直接和间接效应时(图10),农业管理是冬小麦田中蚯蚓丰度的主要驱动因素,其次是跟踪器基面积。
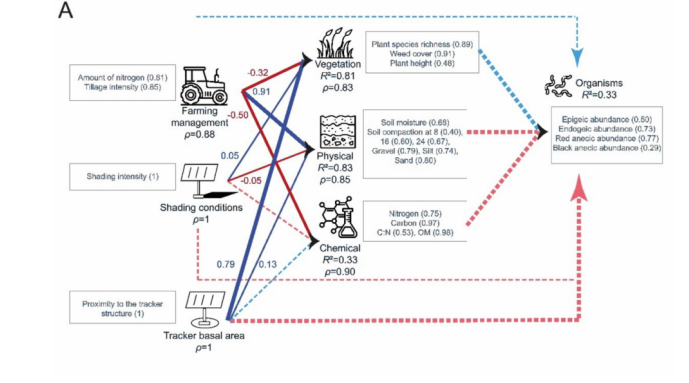
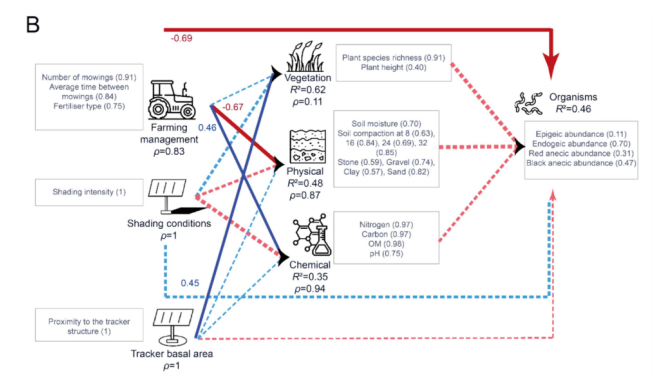
图9. 描述冬小麦田(A)和干草草甸(B)中蚯蚓丰度(“生物”LV)关系的PLS模型:R²是内生潜变量的决定系数,表示由相关外生潜变量解释的方差比例。括号中的值代表交叉载荷。箭头的宽度代表链接的显著性水平。蓝色和红色箭头分别表示正相关和负相关。虚线箭头代表不显著的链接。每个箭头上方的值是外生LV与“生物”LV之间的相关系数(路径效应)。(关于图例中颜色引用的解释,读者请参考本文的网络版本。)
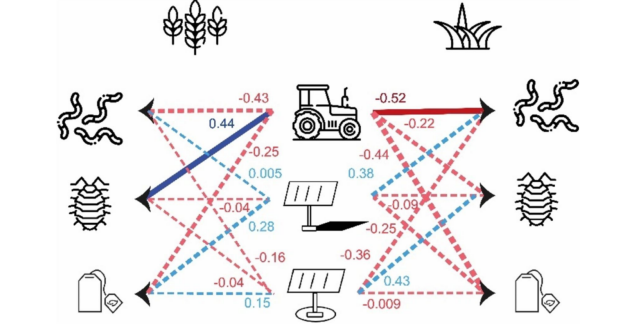
图10. LV跟踪器的总效应(间接和直接)以及LV农场管理对干草草甸和冬小麦田中每种生物(蚯蚓、节肢动物、微生物)LV的总效应的PLS-PM结果。括号中的值代表显变量与其潜变量之间的相关水平。虚线箭头代表外生LV与“生物”LV之间的相关(路径效应)不显著。箭头的大小代表显著性水平。蓝色箭头代表正相关,而橙色和红色箭头代表负相关。(关于图例中颜色引用的解释,读者请参考本文的网络版本。)
干草草甸的PLS-PM模型(图9B)显示,蚯蚓丰度的大部分变异归因于内栖种蚯蚓。内部模型显示,蚯蚓丰度由其潜变量很好地描述(R2 = 0.46)。只有农业管理与蚯蚓丰度呈显著负相关。除植被(ρ = 0.11)外,所有反映型LV都由其MV非常好地描述(ρ > 0.83)。植被由其相关的外生LV很好地描述(R2 = 0.81),主要受到位于跟踪器基面积内的正向影响。土壤物理性质由其相关的外生LV非常好地描述(R2 = 0.83),主要受到农业管理的负向影响。土壤化学性质(R2 = 0.33)由其相关的外生LV很好地描述,主要受到农业管理的正向影响。当同时考虑农业管理和跟踪器相关LV的直接和间接效应时(图12),农业管理是干草草甸中蚯蚓丰度的主要驱动因素,其次是遮荫强度(shading intensity)和跟踪器基面积。
3.2.2.2. 节肢动物活动密度
在冬小麦田中,节肢动物活动密度的大部分变异归因于跳虫(springtails),其次是马陆(millipedes)(图11A)。内部模型显示,节肢动物活动密度由其LV的描述较弱(R2 = 0.24)。除农业管理(正向)和土壤物理性质(负向)分别影响节肢动物活动密度外,所有LV均与节肢动物活动密度无显著相关。所有反映型LV都由其MV非常好地描述(ρ > 0.60)。植被由其相关的外生LV非常好地解释(R2 = 0.83),主要受到跟踪器基面积的正向影响,其次是农业管理和遮荫条件(shading conditions)。土壤物理性质由其相关的外生LV较好地描述(R2 = 0.39),主要受到农业管理的负向影响,其次是跟踪器基面积。土壤化学性质由其相关的外生LV较好地描述(R2 = 0.46),主要受到农业管理的正向影响,其次是跟踪器基面积,但受到遮荫条件的负向影响。当同时考虑农业管理和跟踪器相关LV的直接和间接效应时(图10),农业管理是冬小麦田中节肢动物活动密度的主要驱动因素。
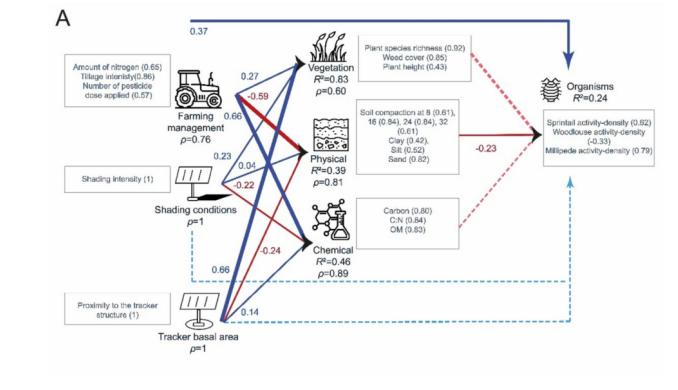
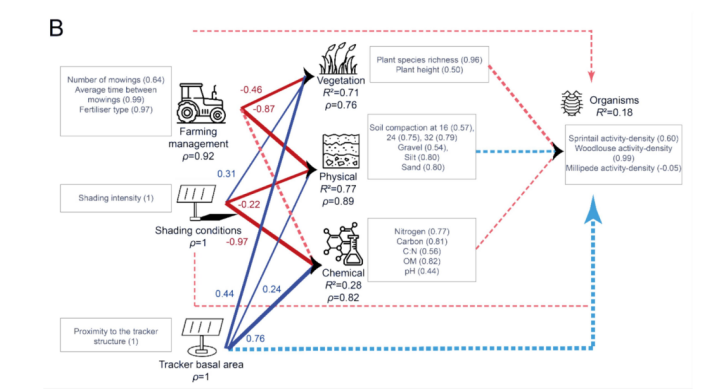
图11. 描述冬小麦田(A)和干草草甸(B)中节肢动物活动密度(“生物”LV)关系的PLS模型:R²是内生潜变量的决定系数,表示由相关外生潜变量解释的方差比例。括号中的值代表交叉载荷。箭头的宽度代表链接的显著性水平。蓝色和红色箭头分别表示正相关和负相关。虚线箭头代表不显著的链接。每个箭头上方的值是外生LV与“生物”LV之间的相关系数(路径效应)。
在干草草甸中,节肢动物活动密度的大部分变异归因于潮虫(woodlouse)(图11B)。内部模型显示,节肢动物活动密度由其LV的描述较差(R2 = 0.18),没有任何LV与节肢动物活动密度显著相关。所有反映型LV都由其MV较好地描述(ρ > 0.60)。植被由其相关的外生LV较好地解释(R2 = 0.71),受到跟踪器基面积和遮荫强度的正向影响,以及农业管理的负向影响。土壤物理性质由其外生LV非常好地描述(R2 = 0.77),主要受到农业管理的负向影响。土壤化学性质由其相关的外生LV解释较弱,主要受到遮荫强度的负向影响,但受到农业管理和跟踪器基面积的正向影响。当同时考虑农业管理和跟踪器相关LV的直接和间接效应时(图10),跟踪器基面积是干草草甸中节肢动物活动密度的主要驱动因素,其次是农业管理。
3.2.2.3. 微生物活性
在冬小麦田中,微生物活性的变异由两个指标同等解释(图12A)。内部模型显示,微生物活性由其LV很好地描述(R2 = 0.32),但没有LV与微生物活性显著相关。所有反映型LV都由其MV非常好地描述(ρ > 0.76)。植被由其相关的外生LV非常好地解释(R2 = 0.67),主要受到跟踪器基面积和农业管理的正向影响。土壤物理性质由其相关的外生LV较好地解释(R2 = 0.47),主要受到农业管理和遮荫条件的负向影响。土壤化学性质由其相关的外生LV较好地解释(R2 = 0.37),受到跟踪器基面积的正向影响,以及遮荫条件和农业管理的负向影响。当同时考虑农业管理和跟踪器相关LV的直接和间接效应时(图10),遮荫强度是冬小麦中微生物活性的主要驱动因素,其次是农业管理和跟踪器基面积。
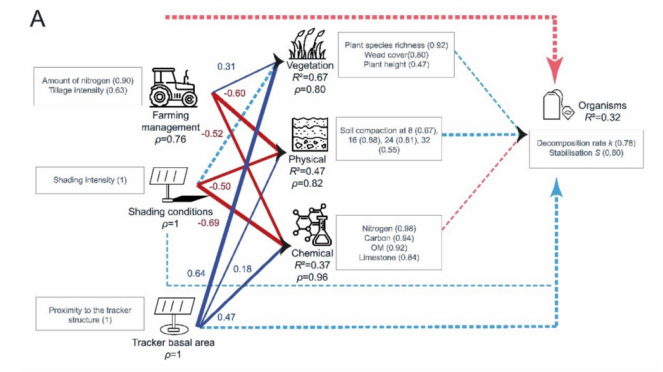
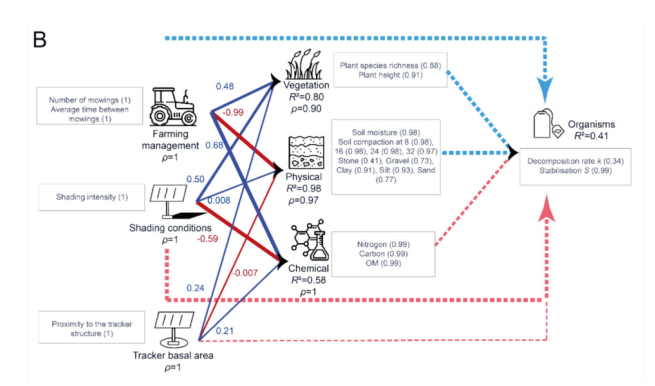
图12. 描述冬小麦田(A)和干草草甸(B)中微生物活性(“生物”LV)关系的PLS模型:R²是内生潜变量的决定系数,表示由相关外生潜变量解释的方差比例。括号中的值代表交叉载荷。箭头的宽度代表链接的显著性水平。蓝色和红色箭头分别表示正相关和负相关。虚线箭头代表不显著的链接。每个箭头上方的值是外生LV与“生物”LV之间的相关系数(路径效应)。
在干草草甸中,微生物活性的大部分变异归因于稳定性指标(stabilisation indicator)(图12B)。内部模型显示,微生物活性由其LV相当好地描述(R2 = 0.41),但没有LV与微生物活性显著相关。所有反映型LV都由其MV非常好地描述(ρ > 0.90)。植被由其相关的外生LV非常好地解释(R2 = 0.80),受到所有这些MV的正向影响。土壤物理性质由其相关的外生LV非常好地解释(R2 = 0.98),主要受到农业管理的负向影响。土壤化学性质由其相关的外生LV较好地解释(R2 = 0.58),受到农业管理和跟踪器基面积的正向影响,以及遮荫条件的负向影响。当同时考虑农业管理和跟踪器相关LV的直接和间接效应时(图10),农业管理是微生物活性的主要驱动因素,其次是遮荫条件。
4. 讨论
据我们所知,这项研究是首批探讨农业光伏(agrivoltaic)PV结构对土壤生物(尤其包括蚯蚓)影响的研究之一(Lambert et al., 2023, Menta et al., 2023)。它将先前关于草地的研究(Bai et al., 2022, Choi et al., 2023, Hassanpour Adeh et al., 2018, Vervloesem et al., 2022)扩展到一年生农田,在我们的案例中是冬小麦田。我们关注负责土壤有机质循环的分类群,以评估太阳能跟踪器的存在是否会相对于农业实践对土壤生物多样性施加额外的限制,或者相反地创造更有利的条件。我们的结果清楚地表明,太阳能跟踪器确实影响环境条件(即土壤的物理和化学性质以及植被特征),其强度取决于与跟踪器的距离,这进而影响土壤生物多样性。然而,无论考虑何种生物类群和土地覆盖类型,农业实践仍然是生物多样性格局的主要决定因素,其影响超过太阳能跟踪器。
4.1. 太阳能跟踪器对环境条件的影响具有距离依赖性
正如预期的那样,距离太阳能跟踪器越近,土壤生物所经历的地面环境条件的改变就越强。首先,在跟踪器基座及其北侧,遮荫强度远高于其他任何地方。减少的太阳辐射与改变的小气候条件有关,特别是降低的温度(例如,Suuronen et al., 2017, Weselek et al., 2021, Yue et al., 2021)。我们自己在四个冬小麦田使用分布在所有方向、距太阳能跟踪器5至35米的40个TMS-4温度记录仪进行的测量证实,靠近跟踪器处(特别是在北侧1米和8米处)的土壤和空气温度显著较低,土壤湿度较高,而与35米处相比。然而,观察到的差异与其他研究相比很小,这些研究大多涉及靠近地面的光伏板阵列,而我们研究的是位于7米高桅杆顶部的点状光伏跟踪器。因此,这些农业光伏装置对局部风速和温度的影响较弱并不令人惊讶(Armstrong et al., 2016, Moscatelli et al., 2022)。其次,无论是在小麦田还是干草草甸中,跟踪器桅杆基部的植物物种丰富度都高于其他地方。这一结果与文献中常见的发现(Bai et al., 2022)形成对比,但再次可能由所研究系统的特定结构解释,因为这些跟踪器是点状的人工基础设施,其对土壤的占用并不比孤立的树木或电线杆更多。拖拉机避免在靠近(跟踪器的地方)行驶。
4.2 农业实践对生物多样性的影响远超太阳能追踪器
通过偏最小二乘路径模型(PLS-PM)分析,我们成功构建了环境变量与土壤生物多样性之间的复杂关系模型。总体而言,农业管理实践是解释土壤生物多样性的最重要因素,而太阳能追踪器的影响十分有限——当综合考虑两类变量对其他变量的多重影响时,其影响甚至不显著(图9、图10、图11、图12)。
农业实践对土壤生物多样性的重要性已广为人知。在考虑农业实践的直接与间接影响后,仅见冬麦田中的节肢动物受到显著正向影响。在一年生作物中,土壤耕作强度(即耕作深度及是否翻耕)已被多次证明对蚯蚓(Capowiez等,2009;Kladivko,2001)、地表节肢动物(Coleman等,2017;Stinner和House,1990)及微生物活性(Mathew等,2012)均存在直接(如直接杀死生物)与间接(如剧烈改变土壤含氧量或湿度条件)的不利影响。肥料类型与用量同样会影响研究生物所需食物的资源性质与可用性(Bünemann等,2006;Geisseler和Scow,2014;Li等,2013)。农业土壤中蚯蚓的活动强度高度依赖于耕作、残留物输入、粪肥施用、化肥及农药使用等管理实践(Mackay和Kladivko,1985;Marinissen和De Ruiter,1993)。对节肢动物而言,耕作可将作物残体混入土壤,从而增加有机质可用性并刺激其种群增长。实际上,停止土壤耕作可能导致跳虫种群减少(Filser等,2002)。通过泥浆或矿物肥料输入的氮素也能促进腐食性大型动物的繁殖(Ponge等,2013)。在干草草甸中,仅蚯蚓受到农业管理强度(即割草强度)的显著负面影响。研究表明割草会减少某些蚯蚓物种(Frazão等,2017),生物量输出会改变土壤条件及可分解有机质的可用性(Morris,1978)。
追踪器下方的植被在冬麦田和干草草甸中均表现出更高的植物物种丰富度:冬麦田的地表覆盖率更高,而草甸的植被高度更大。跳虫受植物生物量(特别是细根数量和根系分泌物)的影响,这些因素会改变非生物土壤条件及水分和养分的可用性(Perez等,2013)。此外,木虱也对植物群落组成敏感,因为这会影响有机质性质及植被物理结构(David等,1999)。由于农民不在追踪器下方耕作,杂草得以生长并为土壤生物提供庇护所,其作用类似于农民和土地开发者自愿种植的绿色或农业生态基础设施(如树篱、休耕地、草带、花坛)。这也正是建议在光伏设施周围保留未耕作或开花空间(例如Grodsky等(2023)针对蚂蚁的研究),或将此类区域用作休耕地以满足欧盟共同农业政策(CAP)要求的理论依据。尽管对植被存在这些影响,我们的研究中并未观察到土壤生物丰度的变化。这可能是因为现有植被类型未能提供额外资源。由此证明在追踪器下方播种开花植物将有利于多种节肢动物。然而,土壤生物质量高度依赖于系统管理,这会直接影响植物(Armstrong等,2016)和土壤无脊椎动物(如通过放牧;Menta等,2023)的多样性。农业管理还可通过植被恢复(Hernandez等,2019)或创新场地管理实践(如光伏设备与多年生本土植被的协同布局)促进碳封存(Choi等,2023),从而改善光伏装置的物理和微生物质量。
4.3 结论
本研究强调,与农业实践相比,双轴太阳能追踪器对土壤环境和生物多样性的影响微乎其微。但需要承认的是,本研究涉及的光伏结构并非典型配置——它采用离散式高位垂直架构,而非文献中主要关注的低位水平固定面板阵列。我们明确证实太阳能追踪器主要通过遮荫和 tracker 邻近区域的土壤属性对环境条件产生微弱影响。由于所研究的太阳能追踪器在调查前已安装2至7年,其安装对土壤特性的影响可能已随时间消退。研究还发现这些影响在草甸和麦田中存在细微差异。未来需要在更广泛的土壤类型和生物气候区域,针对多种作物(如谷物、油籽作物、甜菜、马铃薯)和草地类型(如干草草甸、牧场)开展更多研究,以进一步完善相关影响的评估。本研究中的基础设施因高度较大(面板距地7米)导致阴影区域移动较快(Graham等,2021;Noirot-Cosson等,2022;Suuronen等,2017),下方田地仅受到临时性遮荫。因此有必要评估其他可能对土壤功能影响更大的光伏结构类型。但这些初步结果表明,离散式太阳能追踪器能有效实现作物/牧草与能源生产的土地共享,同时最大限度降低对当地环境及土壤生物多样性的影响,符合可持续发展目标。