


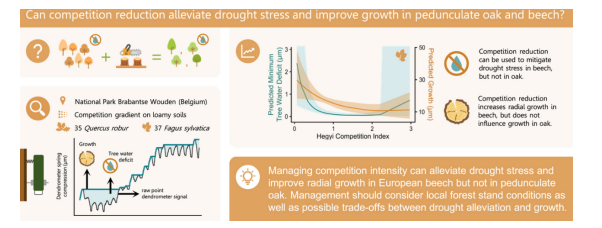
缓解欧洲水青冈和夏栎的干旱胁迫:竞争调控作用
发布时间:2025-09-15 10:52 | 点击次数:92
缓解欧洲水青冈和夏栎的干旱胁迫:竞争调控作用
本文由上海赛弗生物科技有限公司翻译,旨在为相关科研人员提供科学参考,由于我司水平有限,错漏之处在所难免,敬请谅解指正!
原文题目:Mitigating drought stress in European beech and pedunculate oak: The role of competition reduction

亮点
• 研究了干旱胁迫和生长与竞争强度的关系。
• 降低竞争强度可缓解或加剧水青冈的干旱胁迫(存在物种特异性响应)。
• 降低竞争强度对水青冈的生长产生积极影响。
• 夏栎的干旱胁迫和生长不受竞争强度影响。
• 竞争调控效应具有物种依赖性。
摘要
干旱对全球森林生态系统构成重大威胁。需要采取因地制宜的有效森林管理策略以减轻干旱的负面影响,其中竞争调控是通过提升水分可利用性来缓解干旱胁迫的重要措施。本研究以欧洲水青冈(Fagus sylvatica)和夏栎(Quercus robur)为对象,在比利时布拉班特伍登国家公园沿竞争强度梯度布设72棵树木的点式树木径向生长记录仪,通过每日最小树木水分亏缺(TWDmin)量化干旱胁迫,并结合日径向生长量分析竞争强度的影响。研究结果显示:
物种特异性响应:
欧洲水青冈:竞争强度降低(1<2时)可缓解干旱胁迫并促进生长,但过度降低(CI<0.5)反而加剧胁迫
夏栎:干旱胁迫与生长在整个竞争梯度中均无显著变化
环境驱动机制:
· 降水通过降低TWD min促进生长
· 饱和水汽压差则引发相反效应
管理启示:
竞争调控并非普适性解决方案。欧洲水青冈存在"生长-抗旱"权衡:低竞争环境(CI<0.5)虽持续促进生长,但会加剧干旱胁迫。建议重启跨区域、多树种、全土类的长期疏伐试验,为不同立地条件下的抗旱管理提供系统支撑。
关键词:点式树木径向生长记录仪,温带森林,森林经营,竞争强度,干旱胁迫,树木水分亏缺,径向生长
Point dendrometers,Temperate forests,Forest management,Competition intensity,Drought stress,Tree water defici,Radial growth
1. 引言
气候变化已成为全球森林面临的重大威胁之一,导致干旱、风倒和虫害等干扰事件频发(Forzieri等,2022;IPCC等,2022)。这些干扰严重影响森林功能与生态系统服务供给。以干旱为例,其通过引发水力衰竭与碳饥饿(Choat等,2018;Sevanto等,2014;Weed等,2013),导致冠层落叶加剧、树木生长衰退,极端情况下甚至造成死亡率上升(Allen等,2015;Choat等,2018;Senf等,2018)。当干旱与高温事件叠加形成"高温干旱"复合事件时(Allen等,2015;Senf等,2020),蒸散作用增强会进一步扩大树木水分需求,加剧干旱胁迫(Allen等,2015;Dale等,2001;IPCC等,2022;Senf等,2020)。未来气候预测表明,由气候变化引发的干旱与热浪事件在频率、持续时间和强度上均将显著增加(IPCC等,2022),这凸显了森林保护工作的紧迫性。
为应对这些挑战,学者提出通过气候智慧型森林经营策略缓解干旱胁迫(Nabuurs等,2017,2018)。其中一种策略是通过提高疏伐强度降低林分密度,从而减少树木间对水分这一干旱期关键限制资源的竞争。Sohn等(2016)的荟萃分析表明,疏伐虽对针阔叶树种的作用机制不同,但总体上能提升树木生长稳定性。针叶树在疏伐后表现出更强的干旱恢复力与韧性(Sohn等,2016),多项研究证实低密度林分中针叶树在个体和林分水平均具有更好的抗旱稳定性(Bottero等,2021;Navarro-Cerrillo等,2023;Schmied等,2022;Steckel等,2020)。Bradford等(2022)甚至发现高密度林分中干旱导致的死亡率更高。
相较于针叶树,阔叶树在疏伐林分中表现出更强的干旱抵抗能力(Sohn等,2016)。Schmitt等(2020)与Weigel等(2023)研究进一步证实疏伐对阔叶树种生长稳定性的积极影响。但Sohn等(2016)的荟萃分析中阔叶树研究数量有限且偏重地中海物种,使疏伐对其他地区阔叶树的抗旱效益存疑。Niccoli等(2020)报道疏伐对夏栎(Quercus robur)的干旱响应影响不一致,Trouvé等(2017)强调无梗花栎(Quercus petraea)中社会 status 的重要性——优势木的生长稳定性似乎不受林分密度影响。Mausolf等(2018)发现欧洲水青冈(Fagus sylvatica)在经营林分中干旱敏感性升高,归因于林内小气候变化(如气温升高、湿度降低)(Leuschner等,2023)及树体结构改变(如树冠扩大增加个体水分需求)(Jump等,2017)。
既有文献强调干旱期立地条件(如土壤类型、气候、树种)对经营策略的关键影响(Castagneri等,2022;Pardos等,2021;Weigel等,2023),这要求制定管理建议需因地制宜。结论的矛盾性与背景依赖性或许可解释为何多数林务员虽意识到气候变化负面影响,却未相应调整经营实践(Sousa-Silva等,2016)。
许多研究通过年径向生长量(如年轮)分析树木干旱响应(Olivar等,2022),但年度尺度可能掩盖短期响应特征。树木径向变化呈昼夜节律:日间蒸腾导致茎干收缩,夜间水分补充引起膨胀(Zweifel等,2021),而细胞分裂与扩大引发的不可逆膨胀即为生长(Zweifel等,2016)。这种收缩-膨胀动态受气温等影响蒸腾的环境因子调控(King等,2013;Oberhuber等,2023)。因此,高时间分辨率的连续监测有助于理解环境变异、干旱等干扰事件与不同树种生长动态的关联(Sánchez-Costa等,2015)。
本研究旨在探究竞争强度(通过不同采伐强度实现)对比利时中部布拉班特伍登国家公园(National Park Brabantse Wouden)古老森林中欧洲水青冈与夏栎的干旱胁迫及生长响应的影响。沿竞争强度梯度选取72株优势木/共优势木,在2022-2023生长季(其中2022年为1991年以来最干热年份)安装点式树木径向生长记录仪。研究聚焦两个问题与假设:(1)降低竞争强度是否会缓解两种树的干旱胁迫(以24小时最小树木水分亏缺TWDmin表征)?假设认为竞争降低将减轻二者TWDmin,但存在效益阈值——超过该阈值后竞争减少的负面效应(如森林气候缓冲能力丧失)将抵消正面效应。相较于喜光的夏栎,顶极演替树种欧洲水青冈偏好湿润小气候,故其效益阈值对应更高竞争水平。两种树种虽均易受持续干旱影响且具有种特异性抗旱策略(van der Werf等,2007;Vanhellemont等,2019),但欧洲水青冈干旱敏感性更高(Meyer等,2020)。(2)竞争降低如何影响两种树的径向生长概率(GROprob)与日生长速率(GROamount)?假设认为竞争减少会提升二者生长指标直至某临界点,超出后不再产生额外增益。例如Gorrod等(2024)发现提高疏伐强度对大径木年直径变化无影响甚至产生负效应。
2. 材料与方法
2.1 研究区与研究期
研究在布拉班特伍登国家公园(约10,000公顷)的梅尔达尔森林(1,255公顷)与索尼亚森林(4,383公顷)(图1)开展。该古老森林群位于比利时中部,具有悠久的经营历史。当前经营多采用QD原则(目标树经营体系),重点管理特定目标树,其余林木保持自然状态。索尼亚森林拥有全球最高的欧洲水青冈,优越立地与气候条件使树高超过45米(Vandekerkhove等,2018),240年以上古树仍保持年均4.75毫米的直径增长。
该地区的气候被归类为温带气候,其特点是冬季温和多雨,夏季凉爽湿润(Koninklijk Meteorologisch Instituut, 2023a)。在1991年至2020年期间,年平均气温和总降水量分别为11°C和837毫米。我们研究期间的2022年和2023年的年平均气温分别为12.2°C和12.1°C,属于自1991年以来的最高记录值(Koninklijk Meteorologisch Instituut, 2023b, Koninklijk Meteorologisch Instituut, 2024)。平均最低气温(2022年:8.1°C,2023年:8.6°C)和最高气温(2022年:16.3°C,2023年:15.8°C)均显著高于各自的长期平均值(最低:7.3°C,最高:14.7°C)。除了显著变暖外,2022年(701.4毫米,148个降水日)和2023年(1,011.4毫米,207个降水日)的降水量分别属于自1991年以来的最低和最高前五名(Koninklijk Meteorologisch Instituut,2023b, Koninklijk Meteorologisch Instituut, 2024)。2022年夏季(19.6°C,110.6毫米)是自1991年以来最干燥的夏季,其气温和降水量与著名的2018年夏季干旱(19.8°C,134.7毫米)相当。
本研究共选择了72株(共)优势的欧洲栎和欧洲山毛榉(图1,表S1和S2)。Meerdaal森林包含25个欧洲栎样地和11个欧洲山毛榉样地,而Sonian森林则有10个欧洲栎样地和26个欧洲山毛榉样地。所选树木均位于以欧洲栎或欧洲山毛榉为主的林分中,且树龄至少为80年。此外,所有树木均生长在排水良好的壤土上,这是这些森林综合体的主要土壤类型。所选树木必须看起来健康,无明显损伤。此外,我们根据最近采伐干预的不同程度,沿着竞争强度梯度选择了树木。该梯度包括最近没有干预的森林保护区、进行单株或多株选择性间伐的区域,以及进行较大规模更新采伐的地点,包括小面积皆伐和群状采伐,无论是否保留树木。这些采伐干预导致了不同程度的树冠释放和邻近树木之间的竞争减少。沿梯度选择的样地的胸高断面积范围为8至57平方米·公顷−1(中位数:23平方米·公顷−1,表S1和S2)。值得注意的是,所有林分在过去都经过管理,包括森林保护区。因此,这些保护区应被视为几十年前停止管理的先前管理林分,目前正经历着林冠闭合的增加(Vandekerkhove等,2021)。
每个样地的林分调查以所选树木为中心进行,遵循佛兰德斯森林调查的协议(Govaere,2019)。记录了所有树木的胸高直径(DBH)和与所选中心树的距离,包括(i)在9米半径内周长为22–122厘米的树木,以及(ii)在18米半径内周长超过122厘米的树木。
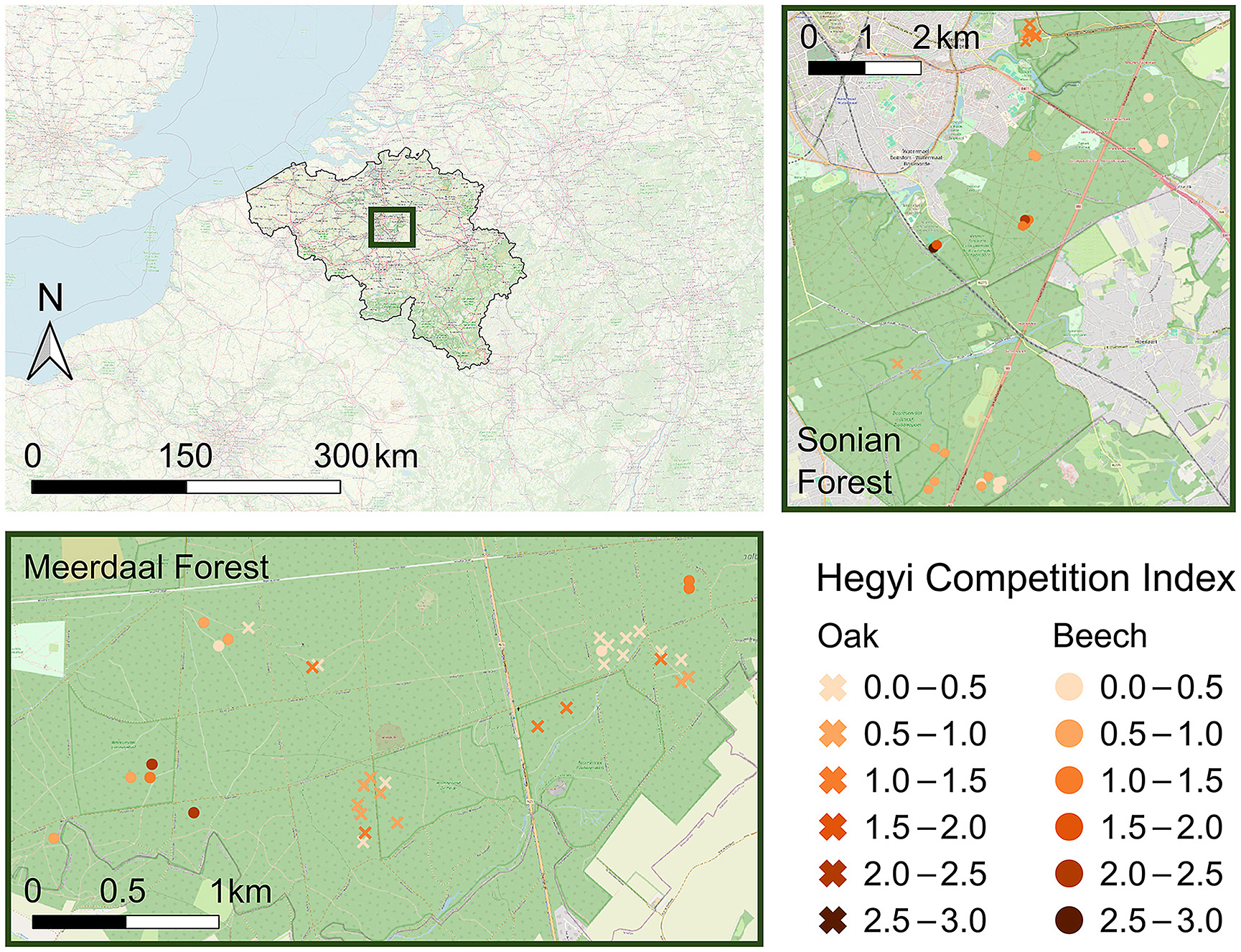
图1. 研究区域概览,显示梅尔达尔森林与索尼亚森林在比利时境内的位置及样地分布。欧洲水青冈(37个样地)与夏栎(35个样地)的颜色梯度表示赫吉竞争指数(Hegyi Competition Index)所表征的竞争强度变化范围,从未经经营管理的森林保护区(高竞争指数)延伸至集约经营林分(低竞争指数)。
2.2 环境数据
每日气象数据,包括最高气温(°C)、相对湿度(%)和降水量(毫米),来自比利时皇家气象研究所的自动化标准化气象站。对于Meerdaal森林,选择了Beauvechain气象站,距离样地4.8至8.6公里。对于Sonian森林,Ukkel气象站作为数据源,距离样地5.4至7.3公里。由于佛兰德斯的地形相对平坦,气象站测量的气候条件与样地相似。
从气象站测量中计算了几个指标。首先,水汽压差(VPD)被纳入,因为除了土壤水分状况外,它是与水分相关的树干半径变化的主要驱动因素(Brinkmann等,2016;Ehrenberger等,2012;Nehemy等,2021)。VPD是根据每日最高气温(Tmax)和相对湿度(RH)计算的,使用August-Roche-Magnus方程(Alduchov和Eskridge,1997)确定每日最大VPD(VPDmax):
(1)![]()
由于数据可用性的限制,无法直接考虑土壤湿度。相反,每日降水量被用作水分可用性的替代指标。此外,降水频率被估计为前两周的雨天数(降水量>1毫米),以描述近期降水事件后土壤中可能的水分储备。
日长(即白昼小时数)使用R包‘insol’根据样地的平均纬度确定(Corripio,2021)。
2.3 森林管理特征
为了描述由采伐干预引起的竞争强度梯度,考虑了两种描述符。首先,选择了Hegyi竞争指数(CI)(Hegyi,1974)来量化中心树所经历的竞争。CI的计算公式为:
(2)![]()
其中,disti是邻居树i与中心树之间的距离(米),DBHi是邻居树i的胸高直径(厘米),DBHcenter tree是中心树的胸高直径(厘米)(图S1)。其次,根据管理记录(如木材销售目录和森林管理计划)确定了样地自上次采伐干预以来的时间(Timeintervention)。
除了CI和Timeintervention外,还计算了每个中心树周围的结构多样性,作为18米样地内竞争树木胸高直径的变异系数(Schmied等,2022)。Schmied等(2022)发现,结构多样性可能通过改善生长和减少压力对中心树有益。
为了考虑年龄对树木生长的影响,中心树的年龄通过计数胸高处取样的树芯的年轮数来确定。当树芯不包含髓心时,根据Rozas(2003)描述的方法,基于从最后一个年轮到血管射线汇合点的距离估计缺失的年轮数。
2.4 点状树木径向生长记录仪Point dendrometer
在2022年和2023年的生长季节(5月至9月),使用TOMST D1点状树木径向生长记录仪(树径仪)(TOMST,2019)记录了树干半径的变化。点装树径仪安装在72个选定中心树的北侧胸高处,以避免阳光直射传感器。在安装前去除树皮的外层死皮,以减少树皮的吸湿膨胀和收缩的影响(Oberhuber等,2020)。测量间隔设置为15分钟,树干收缩和膨胀的分辨率小于1微米(TOMST,2019)。传感器表现出轻微的温度敏感性(补充信息C),这意味着树干半径波动可能由树干水分变化或温度敏感性引起。根据我们现场测量的平均每日温度范围14°C,与温度相关的树干半径波动估计约为7微米(0.46微米·°C−1),而根据制造商TOMST的数据约为2微米(0.12微米·°C−1)。然而,我们并未设定解释的最低阈值,仅建议谨慎,因为春季压缩和温度波动之间的相关性较弱(<0.3,补充信息C)。
使用R版本4.4.2(R Core Team,2024)和‘treenetproc’包版本0.1.6(Knüsel等,2021)处理树径仪数据。工作流程包括几个步骤。首先,排除了错误数据期,包括传感器故障、明显干扰(如破坏、野生动物)以及(重新)安装后的最初两周,以允许传感器稳定。另一种错误数据形式是由于20棵树的生长在第二个生长季节末超过了传感器的测量范围(最大弹簧压缩:8,890微米)。一旦弹簧压缩超过8,800微米,所有后续测量数据均从数据集中删除。其次,将数据在时间上对齐到固定的15分钟间隔,以考虑信号中的较小时间偏移。第三,通过调整每个传感器的proc_dendro_L2函数的tol_jump和tol_out参数,执行异常值和跳跃去除,以优化信号(表S1和S2)。
接下来,通过零增长概念(Zweifel等,2016)从信号中提取径向生长(GRO)和树木水分亏缺(TWD)。该概念指出,树干收缩期意味着没有增长,树径仪信号超过先前最大值的每次增加被视为增长(图2)。低于先前最大值的变化表明TWD,计算为零增长线与信号之间的差值。从TWD中确定每日最小TWD(TWDmin)(图2)。如果TWDmin大于零,表明夜间水分储备未完全补充,表明水分供应不足,从而产生干旱压力和抑制生长过程。从径向生长中确定每日径向生长量(GROdaily)(图2)。如Zweifel等(2016)所述,零增长概念被证明是分离与水分储存动态相关的树干半径变化的有效假设。然而,关于增长的确切时间存在一些不确定性,这些不确定性随着整合期长度的增加而平衡。在一个15分钟间隔内低估的增长增量被后续15分钟间隔内的高估所抵消。在较长时间(周、月或年)的总和中,时间不准确性不再起作用。在本工作中,分析的最小时间分辨率为每日。
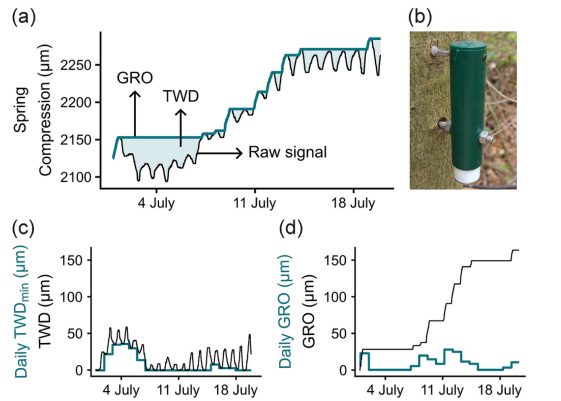
图2.(a) 2022年7月一株有柄栎树木质部径向变化原始信号示例。根据零生长概念(Zweifel等,2016)确定了树木水分亏缺(TWD)和径向生长量(GRO)。(b) TOMST点式树木径向变化测量仪安装于欧洲水青冈树干的情形。(c) 通过零生长概念从原始信号中提取TWD,并转换为日最小TWD值。(d) 根据树木累积GRO数据提取每日GRO增量。
2.5 统计分析
本研究采用‘glmmTMB’程序包(Brooks等,2017)构建广义线性混合模型(GLMMs)。响应变量(TWDmin、GROdaily)呈零膨胀伽马分布,需采用广义模型进行拟合。针对TWDmin,选择Tweedie分布处理零值问题;针对GROdaily,采用伽马栏栅模型进行两阶段分析:首先通过逻辑回归建立径向生长概率模型(GROprob),其次对存在生长的天数(GROdaily > 0),使用伽马分布对实际观测生长值(GROamount)进行建模。具体拟合模型如下:

建模前对所有数值型解释变量进行标准化处理,各变量的选取依据见表S3。需要说明的是,模型引入了树木测量仪编号(Dendrometer ID)作为随机效应以处理数据的层次结构,同时采用一阶自回归(AR1)结构消除时间自相关性。由于森林经营干预措施针对研究区内具有发展潜力的单木,本研究所有分析均基于树木尺度而非林分尺度。
3 结果
3.1 树木水分亏缺
在2022和2023年生长季期间,欧洲栎和欧洲水青冈均表现出较高的最小树干收缩值(TWDmin)(图3)。TWDmin存在显著的年内和种内变异:欧洲栎中位TWDmin为9微米(四分位距:2-43微米,最大值:339.5微米),欧洲水青冈中位TWDmin为7微米(四分位距:2-27微米,最大值:142.2微米)。两种树种在2022年生长季初期及夏季均经历了干旱胁迫期(TWDmin > 0)。需要特别说明的是,由于TWD受树干形态和树皮解剖结构影响显著(Brinkmann等,2016;Peters等,2023),除非经过标准化处理,否则不应直接比较不同树种间的绝对TWDmin值。
根据TWDmin模型分析结果,Hegyi竞争指数(CI)和干预时间(Timeintervention)对欧洲水青冈的树木水分亏缺(TWDmin)具有最显著影响(图4;表S4和S5),而欧洲栓皮栎的TWDmin则不受CI或Timeintervention影响。欧洲水青冈的TWDmin在CI值介于1-2时达到最低值。若考虑不确定性区间范围,当CI较高时(不确定性较大),显著性趋势会有所减弱(图S3)。结构多样性对TWDmin无显著影响。环境变量对TWDmin的预测贡献相对较小,但在两个树种中呈现相似变化趋势:随着饱和水汽压差(VPD)升高,TWDmin相应增加,但该效应在昼长较长时会减弱(图S4);降水量增加则会导致TWDmin降低。
3.2 生长特征
欧洲栓皮栎与欧洲水青冈的日生长量(GROdaily)存在显著的年内变异和种内个体差异(图3)。在发生生长的日期(GROdaily > 0),欧洲栓皮栎的日生长量中位数为22.8微米(四分位距:4-32微米,最大值:440微米),欧洲水青冈为25.8微米(四分位距:4-38微米,最大值:255微米)。2022年两种树种的平均累积生长曲线均高于2023年(图3f与3g),这可能表明干旱遗留效应的影响。
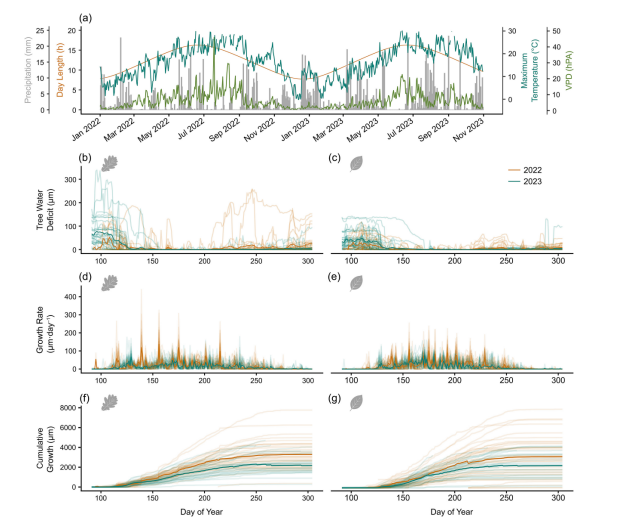
图3.(a) 2022年与2023年多项气象特征,包括最高气温、饱和水汽压差(VPD)、降水量及昼长。(b–g) 2022和2023年生长季期间,欧洲栓皮栎(左)与欧洲山毛榉(右)的树木水分亏缺(TWDmin)、生长速率及累积生长量。粗线表示所有监测树木的平均响应(背景细线为个体数据)。
3.3 生长量模型分析
GROamount的模型趋势与GROprob模型具有可比性。欧洲栓皮栎的生长量未受管理措施影响,而欧洲水青冈的生长量随竞争强度增加显著降低,随结构多样性增加而上升。与GROprob类似,树龄对欧洲水青冈的径向生长产生负向影响(图5)。昼长是决定GROamount的最主要影响因素(图4):较长昼长条件下生长量增加,而高饱和水汽压差(VPD)会限制生长量(图S6)。降水量增加促进GROamount提升,但降雨频次增加反而会降低生长量(图S6)。
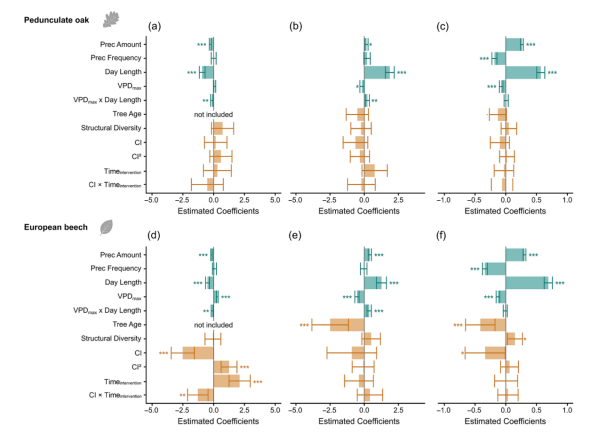
图4. 欧洲栓皮栎与欧洲水青冈的TWDmin(a,d)、GROprob(b,e)和GROamount(c,f)标准化解释变量拟合模型的系数估计值。各变量均标注统计显著性及不确定性,其中蓝色解释变量为非生物环境因子,橙色解释变量为可通过伐木干预改变的变量。欧洲栓皮栎和欧洲水青冈的详细系数估计值、标准误及P值分别见表S4和表S5。(注:Prec为降水量英文缩写)
4 讨论
4.1 森林管理对TWDmin、GROprob和GROamount的影响
本研究旨在探究采伐干预产生的竞争强度对欧洲水青冈和欧洲栓皮栎干旱响应与生长的影响。第一假设认为竞争减弱将缓解两种树种的干旱胁迫,且存在一个预期阈值,超过该值后竞争减少的负面效应将超过正面效应。第二假设假定竞争强度降低会促进两种树种的日径向生长,但这种促进效应会逐渐饱和,进一步降低竞争不会带来额外收益。
研究结果未强烈支持第一假设。竞争效应和末次疏伐时间的影响具有物种特异性:欧洲栓皮栎的竞争减少未显著改变干旱胁迫,这与Trouvé等(2017)关于无梗花栎生长稳定性与林分密度无关的发现一致;而欧洲水青冈的竞争强度显著影响干旱胁迫(图5)。当CI低于0.5时,其TWDmin上升,且末次疏伐后时间越长该效应越强。强度管理似乎加剧干旱胁迫,这与Mausolf等(2018)在德国北部发现的管理林分中水青冈干旱敏感性更高的结论一致。另一方面,高竞争水平(CI>2)也会导致干旱胁迫加剧,与Cescatti和Piutti(1998)关于强竞争下水青冈对水分平衡敏感性增加的报道相符。需注意的是,高CI范围仅有一个样地代表,导致置信区间扩大,因此CI>2时TWDmin增加的可信度存疑。欧洲水青冈存在最小化干旱胁迫的最优林分密度(1<2),但高竞争强度的可解释性有限。
关于第二假设,欧洲栓皮栎的生长量未受管理措施显著影响(表S4),假设被拒绝。Niccoli等(2020)发现管理效应因处理强度和类型而异,仅高强度择伐(75%断面积削减)促进生长。欧洲水青冈的径向生长随竞争减少持续改善(图5;表S5),Mausolf等(2018)也发现管理林分中生长量更高。林分水平上降低密度同样促进生长(Bosela等,2016;Weigel等,2023),但强度疏伐会降低林分生长(Bosela等,2016)。
基于上述发现,欧洲水青冈需在缓解干旱胁迫与促进个体生长间寻求平衡(类似Mausolf等2018的权衡关系)。应避免强度竞争削减(因其加剧干旱胁迫,且随时间推移效应增强),这可能源于森林小气候缓冲能力丧失(Mausolf等,2018;Weigel等,2023)。闭冠层和高结构多样性林分具有更稳定的小气候,可缓解大气候波动(de Frenne等,2021)。这也可解释结构多样性对欧洲水青冈生长的正向效应(与Schmied等2022关于挪威云杉的发现一致)。建议对欧洲水青冈实施轻度至中度疏伐,同时保持高结构多样性。应避免极开阔或极密集林分,因其分别加剧干旱胁迫和抑制径向生长。
研究结果表明物种对管理的响应存在差异:欧洲水青冈受益于林分密度管理,而欧洲栓皮栎无显著响应。这种物种特异性响应与文献中关于物种对环境因子(Etzold等,2022;King等,2013;Schäfer等,2018)和管理强度(Juodvalkis等,2005;Sohn等,2016;Vernon等,2018)差异化响应的记载一致。此外还有两种解释:其一,欧洲栓皮栎的竞争强度梯度较窄(尤其高竞争端),其断面积变幅也较小(表S1、S2)。作为喜光树种,栓皮栎林分结构更开阔,竞争梯度相对有限;其二,欧洲水青冈因解剖结构特性(Meyer等,2020;Scharnweber等,2011)对干旱更敏感——散孔材比环孔材(栓皮栎)具有更高气候敏感性(Elliott等,2015),且细根主要分布在0-30厘米土层(Leuschner,2020),而栓皮栎深直根可获取深层土壤水分(Eaton等,2016)。
需指出本研究仅涉及一个显著干旱年(2022年),但该年干旱程度不容忽视:尽管2021年是1991年以来最湿润年份之一,且壤土持水性优异,但30米深的地下水位使树木完全依赖土壤储水,而2022年夏季是1991年以来最干旱季节,土壤储水不足以缓冲干旱影响。
4.2 环境条件对最小树干水分亏缺、生长概率及生长量的影响
除经营管理外,日长、饱和水汽压差(VPD)及降水量也是关键变量。欧洲水青冈和有柄栎对这些变量的响应在显著性和影响程度上表现相似。
日长是影响干旱胁迫和树木生长的最重要环境因子(图4)。生长季(5-9月)内白昼较短时,树木干旱胁迫程度最高。随着白昼延长,干旱胁迫减轻,而形成层活动和径向生长速率随光周期增加,促使树木生长加速。夏至前后(6月21日,年积日172天)达到生长概率和生长量的峰值,此时树木生长进入高峰期(Etzold等,2022;Rathgeber等,2016;Rossi等,2006)。已知日长(光周期)是重要的生长约束因子,它为树木提供了生长窗口期,既能避免晚霜危害(Vitasse和Basler,2013),又能通过完成次生细胞壁木质化为越冬做好充分准备(Rossi等,2006)。
在此窗口期内,天气条件将制约生长并影响干旱胁迫(Etzold等,2022;King等,2013;Schäfer等,2018)。本研究发现较高的VPD会加剧干旱胁迫并抑制生长(图S4-S6)。高VPD条件常导致植物气孔导度和光合作用减弱,进而增加蒸腾耗水并可能引发碳饥饿,这与多项研究中观察到的生长衰退现象一致(Babst等,2019;Grossiord等,2020;Choat等,2018;King等,2013;Sevanto等,2014)。
与Duchesne和Houle(2011)及Etzold等(2022)的研究相似,降水特征的影响相较于其他解释变量相对有限(图4)。强降雨事件可缓解干旱胁迫并促进生长,这与现有文献结论一致(Mausolf等,2018;Navarro-Cerrillo等,2019),其主要机制是提高了土壤水分有效性(Deslauriers等,2003;Etzold等,2022;King等,2013)。除通过土壤水分影响最小树干水分亏缺外,降雨还能通过降低VPD来缓解干旱胁迫(Zweifel等,2005)。但过于频繁的降雨会降低生长量,这种现象在水分充足时也可能出现。
4.3 对森林经营管理的启示
气候变化背景下,VPD预计将持续上升,降水模式将更趋不规则且难以预测,导致干旱事件更频繁剧烈(Allen等,2015;IPCC等,2022)。间伐作为缓解水资源竞争的措施被广泛推荐。然而本研究显示,在现有竞争强度梯度、气候和土壤类型条件下,有柄栎通过减轻竞争来缓解干旱胁迫或促进生长的效果并不显著。对于欧洲水青冈,竞争强度确实会影响单木水平的干旱胁迫和生长:适度减轻竞争可初步缓解干旱胁迫,但过度降低竞争强度(竞争指数<0.5)反而会加剧胁迫;而径向生长始终受益于竞争强度降低。这表明在缓解干旱胁迫与促进生长之间存在权衡关系。基于此,我们建议对温带壤土林分的欧洲水青冈实施轻度至中度间伐。现有结果尚不足以制定针对有柄栎的干旱胁迫缓解方案(其林分密度范围为9.9–41.5 m²·ha⁻¹)。
这些结果表明,通过降低竞争来缓解干旱胁迫的潜力在该研究地区和树种中具有局限性和物种特异性。本研究强调需综合考虑当地林分条件,避免制定跨物种、跨区域的通用经营方案。未来研究应涵盖更多土壤类型、气候条件和树种。建议基于气候智慧型森林经营理念,重新审视和激活长期间伐试验,从而更深入揭示竞争调控的影响机制。
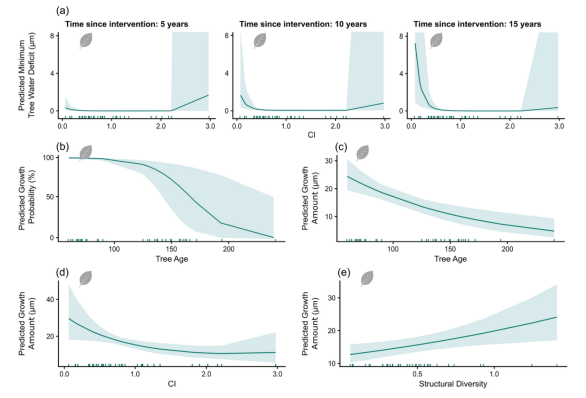
图5. 所有模型中受伐木干预影响变量的偏依赖图。(a)竞争指数(CI)与干预时间(Timeintervention)对欧洲水青冈TWDmin的交互效应。(b-c)树龄对欧洲水青冈GROprob和GROamount的影响。(d-e)竞争指数(CI)与结构多样性分别对欧洲水青冈GROamount的影响。
5 结论
本研究发现以竞争强度表征的经营管理措施会加剧低竞争强度下欧洲水青冈的干旱胁迫,但其生长持续受益于竞争减弱;而有柄栎的干旱响应和生长均未受经营管理影响。这表明降低竞争并非总能缓解或加剧干旱胁迫。对于欧洲水青冈,需在缓解干旱胁迫与促进生长之间寻求平衡,因此应避免高强度间伐。环境变量对干旱胁迫和生长具有持续影响,凸显了其在森林动态中的关键作用,也印证了气候变化对森林健康的重要影响。
需通过扩展竞争强度梯度(尤其对有柄栎)和延长研究周期以涵盖更多干旱事件,进一步探究竞争强度效应。未来应围绕更广泛的干旱条件、树种和土壤类型,深入研究降低竞争作为缓解措施的适用性,从而为优化树木竞争关系、因地制宜制定气候变化应对策略提供科学依据。
附补充材料:
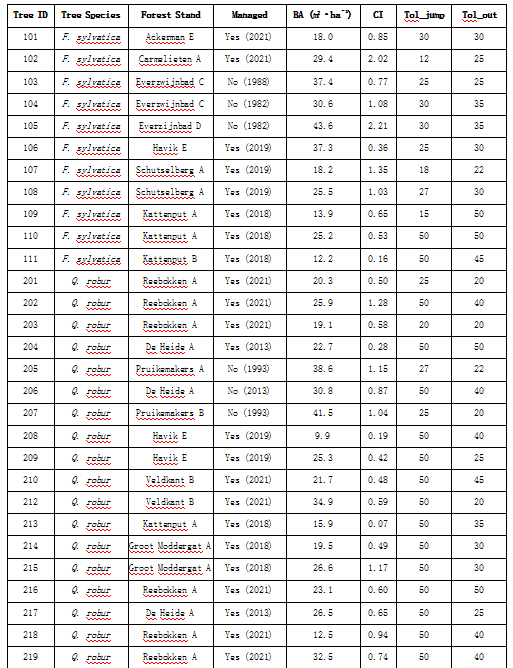
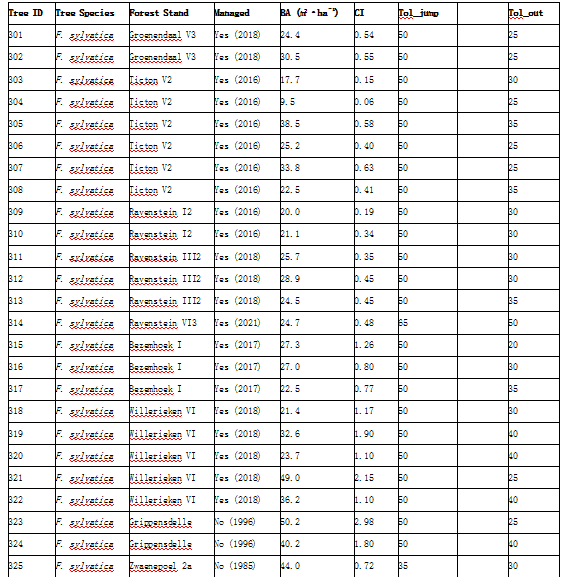
表S1. 该表概述了在Meerdaal森林中选定的树木及其特征。‘Managed’列表示该林分目前是否处于积极管理状态,以及该林分最后一次管理的年份。表中还包括了R包‘treenetproc’算法所使用的tol_jump和tol_out的输入值。请注意,tol_jump和tol_out是输入值,而不是应用的实际阈值。
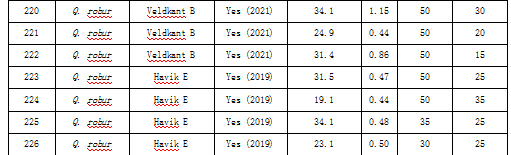
表S2提供了索尼安森林所选树木及其特征的概览。"Managed"(是否管理)一列显示了林分当前是否处于主动管理状态以及最近一次管理的年份。表中还包含了R包"treenetproc"算法所使用的tol_jump和tol_out输入值。请注意,tol_jump和tol_out均为输入参数值,而非实际应用的阈值。
补充信息 B - Hegyi竞争指数
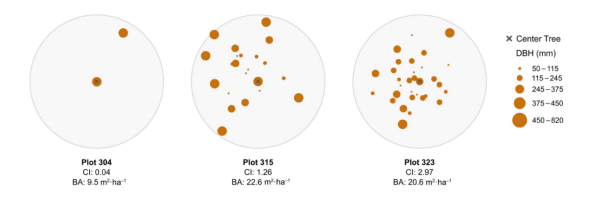
图S1. 三个欧洲山毛榉样地内树木及其胸径(DBH)的可视化分布图,每个样地半径为18米。图中展示了具有最低、中等和最高Hegyi竞争指数(CI)的样地。请注意,基面积(BA)与竞争指数不一定相关。
补充信息 C - TOMST点式树木径向生长仪的温度敏感性
评估树木径向生长仪及其信号质量时,理解传感器的温度敏感性至关重要。热敏传感器可能导致检测信号出现误差或偏差,因为弹簧压缩度的变化可能与温度而非树木生理响应相关。
为评估TOMST点式树木径向生长仪的热敏感性,将传感器安装在派热克斯玻璃板上(图S2)。该玻璃板的热膨胀系数较低(3.3×10–6 °C–1),在观测温度范围(–8.8至37.8°C)内可视为无收缩膨胀现象。传感器不仅记录弹簧压缩度,还通过底部的热电偶监测气温。该装置安装于林内遮阴处以避免阳光直射,并选择昼夜温差最大的样地进行监测。原始生长仪信号显示对温度波动存在微弱逆向响应(图S2a)。这种逆向压缩响应可归因于生长仪不锈钢元件的热膨胀效应:当温度升高时,安装杆的膨胀程度超过弹簧和电位计测量杆,导致生长仪与安装座间距增大;温度下降时则产生相反效应。
根据记录信号,分别计算升温和冷却过程中弹簧压缩度的变化值,再除以与观测位移相关的温差(图S2)。测得升温和冷却阶段的平均热敏感性分别为–0.49 µm·°C–1和–0.43 µm·°C–1,处于Treenet组织(2020)规定的高质量传感器范围内。但需注意,这些测量数据采集时间有限,不能反映传感器的长期稳定性。据制造商TOMST报告,实验室条件下的温度敏感性范围为–0.1至–0.14 µm·°C–1。
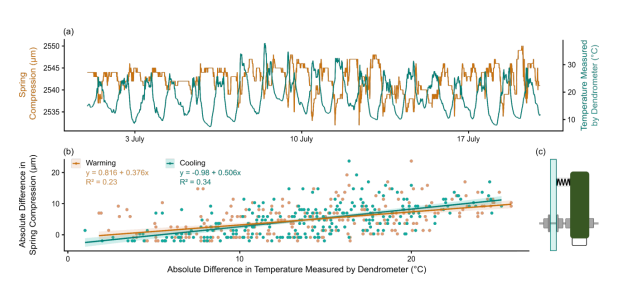
图S2. (a) 2023年7月安装在派热克斯玻璃板上的点式树木径向生长仪的弹簧压缩信号与温度测量示例;(b) 升温期(温度上升阶段)和冷却期(温度下降阶段)弹簧压缩差值与温差的对应关系图;(c) 点式树木径向生长仪在派热克斯玻璃板上的安装示意图。
参考文献
Treenet, 2020. 方法指南 - 点式树木径向生长仪.
补充信息 F- 拟合模型的边际效应图
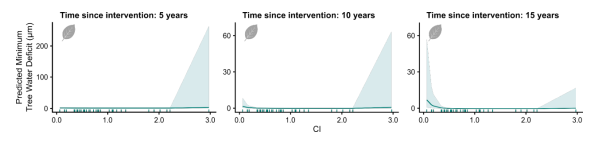
图S3. 干预后时间与CI(冠层开度)交互作用对欧洲水青冈最小树干收缩量(TWDmin)的影响
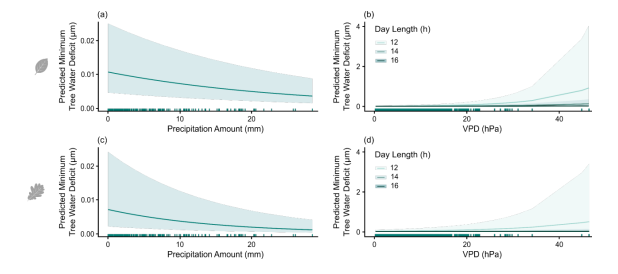
图S4. 基于欧洲水青冈模型(a–b)和欧洲白栎模型(c–d)预测的最小树干收缩量(TWDmin):(a, c)降水量和(b, d)日长与蒸汽压亏缺(VPD)交互作用的影响
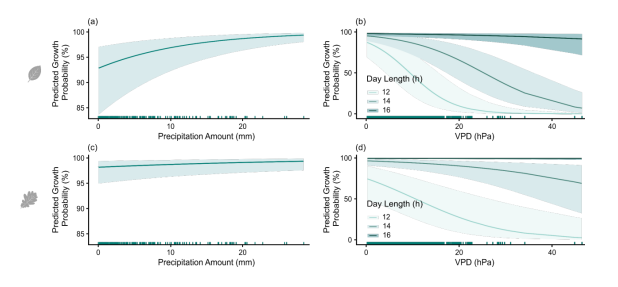
图S5. 基于欧洲水青冈模型(a–b)和欧洲白栎模型(c–d)预测的生长概率(GROprob):(a, c)降水量和(b, d)日长与蒸汽压亏缺(VPD)交互作用的影响
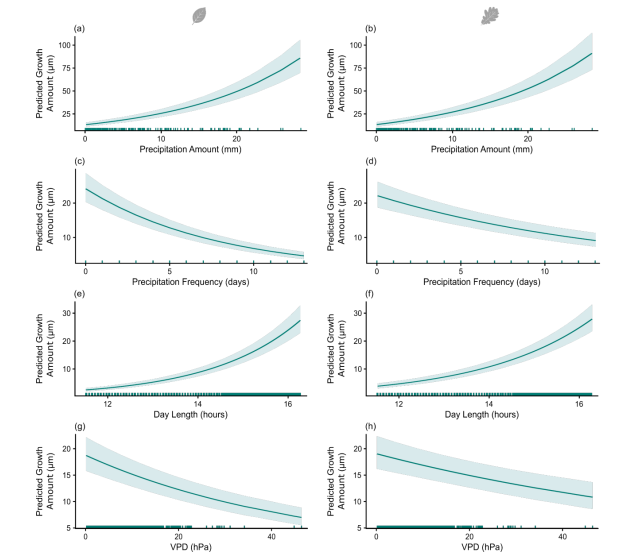
图S6. 基于欧洲水青冈模型(a, c, e, g)和欧洲白栎模型(b, d, f, h)预测的生长量(GROamount):(a–b)降水量,(c–d)降水频率,(e–f)日长,以及(g–h)蒸汽压亏缺(VPD)